
Csak sajnos nem azért, hogy tanuljon, hanem, hogy (úgymond) tanítson.
(2. rész, 3. rész, 4. rész, 5. rész)
(Köszönet a linkért Ambrus Géza Gergelynek.)
 (Szerencsére SexComb nincs vakáción, így egy kis szünet állhat be a poszt-ínséges időkben. - dolphin)
(Szerencsére SexComb nincs vakáción, így egy kis szünet állhat be a poszt-ínséges időkben. - dolphin)
A tervezéselmélet egyik fő érve, amit úton-útfélen előhoznak, az egyszerűsíthetetlen összetettség, ami a hívek szerint egyértelműen bizonyítja az élőlények tervezett voltát.
Maga az érv elég régi, de az ÉRTEM által használt mai formájának alapjait Michael Behe a Darwin fekete doboza című könyvében fektette le, az eredeti meghatározása valahogyan így hangzott:
"A single system which is composed of several interacting parts that contribute to the basic function, and where the removal of any one of the parts causes the system to effectively cease functioning."
"Egységes rendszer, amely számos egymással kölcsönható részegységből áll, amelyek mind hozzájárulnak az alapvető működéshez és ahol a részegységek bármelyikének az eltávolítása a rendszer működésképtelenségét eredményezi."
Az érvelés annyi, hogy ezek a rendszerek nem alakulhattak ki apró változások sorozatával, hiszen minden részegységük szükséges a működésükhöz. "Egyszerűsíthetetlen összetettség nem állhat elő fokozatosan, egy korábbi rendszer jelentéktelen, egymás utáni változásainak eredményeként, mert definíció szerint egy egyszerűsíthetetlenül összetett rendszernek bármely őse működésképtelen."
Behe meg is nevezett pár ilyen rendszert, az eredeti példái közül most a baktériumok ostorával szeretnék részletesebben foglalkozni, mivel magyar nyelven erről írtak eddig a legkevesebbet.
"A baktérium ostora" alatt az Escherichia coli baktérium ostorát értjik általában, amely valóban számos egymással kölcsönható részegységből épül fel, amelyek eltávolítása működésképtelenné teszi az E. coli baktérium ostorát. Ennyiben teljesülnek Behe feltételei, viszont, ha megnézzük, miből is áll a többi baktérium ostora, már merőben más képet kapunk. Szerencsére találtam egy összefoglaló cikket (Pallen M. J. és Matzke N. J. 2006.), így ennek a táblázatát venném át:
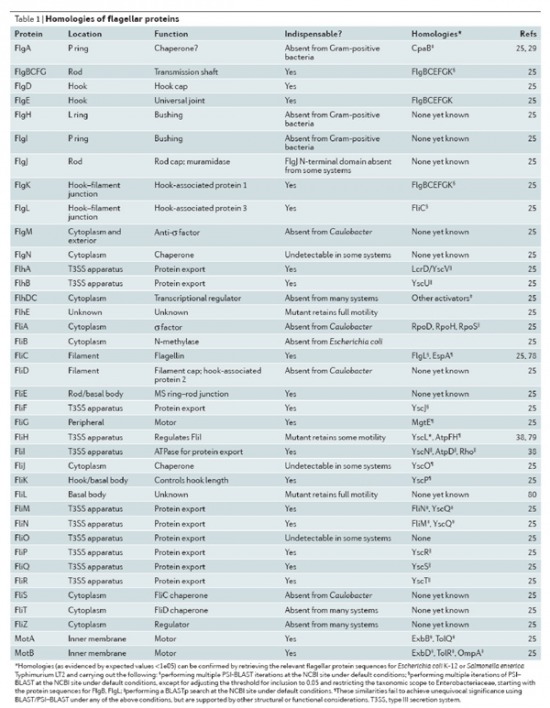
Mit is látunk ezen a táblázaton? Léteznek más baktériumok, amelyek ostora eltérő felépítésű, mint az E. coli-é, ezekből hiányoznak olyan részegységek, amelyek az elsőként vizsgált ostor működéséhez elengedhetetlenek. Sőt, léteznek olyan baktériumok, amelyek ostora az E. coli-énál több alkatrészből áll. Akkor ez hogy van? Mégsem egyszerűsíthetetlenül összetett ez a rendszer? Nézzük meg például az FlgI fehérjét! Ez szükséges az E. coli ostor működéséhez, ha az ezt kódoló gént eltávolítjuk a genomból, az E. coli ostora nem működik (Hizukuri és munkatársai 2006.). Ez eddig egybevág Behe elméletével, ám ha megnézzük, azt találjuk, hogy ez a fehérje hiányzik az összes Gram-pozitív baktériumból. Akkor ez hogy lehet? Van egy egyszerűsíthetetlenül összetett rendszernek egy olyan változata, amelyből hiányzik az egyik alkotórész, mégis működik! Mégiscsak lehetnek működőképes elődei, amelyek nem tartalmazzák minden részegységét, például ott a Gram-pozitív baktériumok ostora! Ugyanígy végigmehetünk a táblázat többi során is és láthatjuk, hogy bizony az "egyszerűsíthetetlenül összetett" rendszer több alkotóeleme is hiányzik működő baktérium ostorokból. Elég egyszerűen sikerült cáfolni ezt az érvelést. Hogy ezt az ellentmondást feloldja, William Dembski kicsit módosította az eredeti meghatározást:
"A system performing a given basic function is irreducibly complex if it includes a set of well-matched, mutually interacting, nonarbitrarily individuated parts such that each part in the set is indispensable to maintaining the system's basic, and therefore original, function. The set of these indispensable parts is known as the irreducible core of the system."
"Egy adott alapvető működést ellátó rendszer egyszerűsíthetetlenül összetett, ha tartalmaz egy készlet egymáshoz illő, egymással kölcsönható, nem tetszőlegesen kiegyénült részegységet, amely készlet minden eleme elengedhetetlen a rendszer alapvető, ezáltal eredeti működéséhez. Ezen elengedhetetlen részegységek a rendszer egyszerűsíthetetlenül összetett magját alkotják."
No most ez a meghatározás, attól eltekintve, hogy jóval ködösebb, mint Behe változata, ebben az esetben csak növeli a homályt, bár tény, hogy jóval megengedőbb, hiszen e maghatározás szerint nem kell az egész rendszernek egyszerűsíthetetlennek lennie, csak egy egyszerűsíthetetlen "magot" kell tartalmaznia. Mondjuk ez még mindig nem magyarázza meg, hogy akkor tulajdonképpen mit is tervezett az értelmes tervező? Mert ugye az E. coli baktérium ostorának az egyszerűsíthetetlen magja nem működik önmagában, kellenek hozzá olyan alkatrészek, amelyek más rendszerekből hiányoznak, tehát nyilván nem egyszerűsíthetetlenül összetettek. Viszont akkor fel kell tételeznünk, hogy az értelmes tervező egy működésképtelen rendszert alkotott, amit evolúciós folyamatok tettek működőképessé. Nem tudom, ez mennyire "értelmes" tervezés.
Létezik egy harmadik lehetőség is, nevezetesen az eredetileg tervezett ostor egy egyszerűbb, ámde működőképes szerkezet volt, amelyhez később kapcsolódtak újabb részegységek, amelyek részben átvették egyes eredeti alkatrészek működését, így ha egy mai ostorból eltávolítjuk őket, az már nem működik nélkülük, de ettől még későbbi toldalékok. Ez a magyarázat tetszetős, ugyanis éppen arra világít rá, hogyan alakulhat ki egy egyszerűsíthetetlenül összetett rendszer lépésenként, evolúciós folyamatok által, ebben az esetben az értelmes tervező már csak fölösleges csökevény az elméletben, ha eltávolítjuk, ugyanúgy megmagyarázza ezen rendszerek létrejöttét.
Ezen kívül is vannak gyenge pontok az elméletben: Az első gond vele, hogy ez esetben egy Gram-pozitív baktérium esetében egy másik rendszerről van szó, mint az E. coli esetében, amely tényt az elmélet egyszerűen nem kezeli sehogyan sem. Ezek szerint "a baktérium ostor" tulajdonképpen nagyon sok különböző, ám egytől-egyig egyszerűsíthetetlenül összetett rendszer gyűjtőneve. Viszont ezt a kérdést én még nem hallottam egyetlen ÉRTEM hívőtől se, illetve nem olvastam egyetlen írásukban sem, ők állandóan "a" baktérium ostorról beszélnek, ami alatt ki tudja hány különböző rendszert értenek, de valamiért kísérletet sem tesznek arra, hogy ezt a bizonytalanságot eloszlassák. Gyakorlatilag kizárólag az Escherichia coli baktérium ostoráról beszélnek, teljes mértékben figyelmen kívül hagyva a tényt, hogy ezen kívül nagyon sok ettől különböző baktériumostor létezik, amelyek felépítése romba dönti az elméletüket.
A másik érdekes része ennek a meghatározásnak, hogy e szerint a rendszer alapvető működése az eredeti feladata is, viszont ez a körkörös érvelés szép példája, hiszen eleve kizárja az esetleges evolúciós változásokat, azaz, amely rendszer egy más működést végző rendszerből alakult ki, máris nem egyszerűsíthetetlenül összetett.
Ha megvizsgáljuk közelebbről ezeket a "részegységeket", amelyekből ez az "egyszerűsíthetetlenül összetett" rendszer áll, megint csak meglepetésben lesz részünk. Mik is ezek a részegységek, amik elengedhetetlenek a működéséhez? Behe professzor biokémikus, ő az egyes fehérjéket tekinti részegységnek. Kiválasztottam hát két fehérjét hasraütéses alapon, egyet a táblázat elejéről, egyet a végéről, legyen mondjuk a FlgD és a MotA.
Mindkettő szándékosan olyan fehérje, amely minden eddig ismert ostorban megtalálható, így mindenképpen egyaránt bele kell hogy essen mindkét "egyszerűsíthetetlen összetettség" meghatározásba. Azt nyilván tudjuk, hogy a fehérjék aminosavakból állnak, így nézzük meg, mi ezen fehérjék aminosavsorrendje egyes élőlényekben!
Hasraütéses alapon kiválasztottam tíz FlgD fehérjét, kijelöltem őket a Uniprot adatbázisában (www.uniprot.org) és megpróbáltam az adatbázis beépített programjának a segítségével egymáshoz illeszteni őket. Az eredmény, azt hiszem megdöbbentő: ezek a fehérjék szinte nem is hasonlítanak egymásra.
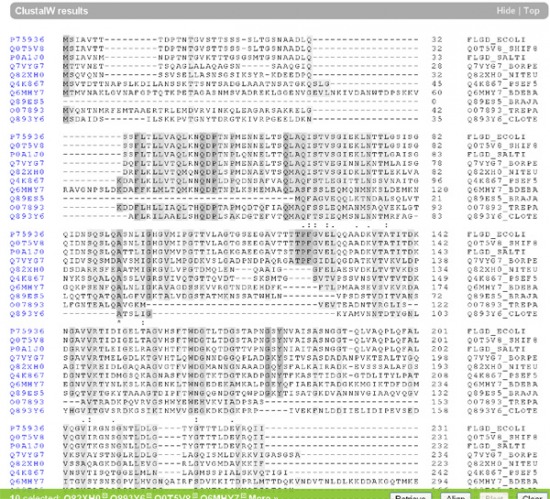
Ha egy megfelelő programmal számszerűsítjük is a különbséget, meglepő eredményeket kapunk:
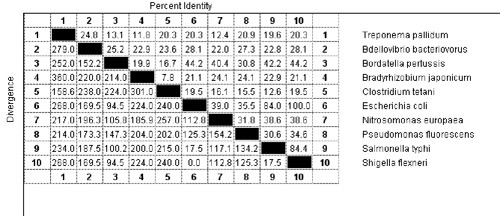
Ha ugyanígy megvizsgáljuk ugyanezen élőlények MotA fehérjéit, hasonló eredményt kapunk:
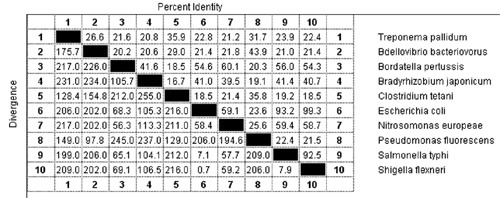
No most ezek a fehérjék egy "egyszerűsíthetetlenül összetett" rendszer alapelemei, amelyek az ÉRTEM elmélet szerint nem jöhettek létre evolúciós folyamatok során, hiszen ezek változásai működésképtelenné tennék az egész rendszert. Ezzel szemben azt látjuk, hogy maguk az alkotóelemek csak mérsékelt hasonlóságot mutatnak. Ha bárki szeretne, nyugodtan keresgélhet még a Uniprot adatai közt, nyilván hasonló képet lát majd. Az ÉRTEM hívei gyakran szoktak értelmes szöveggel példálózni, hogy alátámasszák mondanivalójukat. Hogy érzékeltessem, mit is jelent ez a különbség, fogtam egy ismert szöveget és kicseréltem benne egyes betűket. Elsőként a szöveg betűinek 22% -át hagytam változatlanul, a táblázatban több, hasonló mértékig azonos szekvencia is található, amelyek mind egy-egy működő baktérium ostor részei. Megkérlek, kedves olvasó, találd ki, mi ez a vers!
Eg, ft a hz! tdfgnft, krtd
A rthzests ldrtz gze bgzn?
Ldr, ftre gz he hujmn, hts gd,
Hjkl fgzeacvb h jene ghjklz!
Ugye nem ment! Lássuk, mit mondasz, ha a betűk felét cserélem ki!
Tj, ms a kd! tfghewsó, kgnd
A fzhbjbkn lalil izb brzt?
Lnk, cgjk jr ad iegen, hkt ád,
Hkly fdbvisre a keed dotkwt!
Még mindig elég zavaros. Ha mondjuk csak a betűk húsz százalékát cserélem ki, talán érthetőbb!
Eg, mr a kő! ttúkavyó, kend
A seobtbhn lhkik itt bhnt?
Lkm, ceak jh at ijten, jót ád,
Hohj föuvutte a kind dolgát!
Azért ebből már rá lehet jönni. A megfejtés: Petőfi Sándor: Anyám tyúkja. A feladat nyilván jóval könnyebb, mint egy ismeretlen szöveg megfejtése, mivel az iskolában mindannyiunknak meg kell tanulni ezt a verset, nyilván emlékszünk rá, valamint a szöveg tagolásán, az írásjelek elhelyezkedésén nem változtattam. Mindannyian ismerjük ezt a szövegrészt, mégis teljesen elveszik az információtartalma, ha a szöveget alkotó betűk nyolcvan százalékát lecseréljük. Ellenben ezek az FlgD fehérjék működő baktérium ostorok részei, azaz különbözőségük dacára mégis működnek, annak ellenére, hogy egymásra csak olyan húsz százalékban hasonlítanak. Itt kezdenek szaporodni a kínos kérdések: Tulajdonképpen melyik FlgD fehérjét teremtette az értelmes tervező? Az ÉRTEM hívei valamiért az E. coli baktérium ostorát tűzték zászlajukra, de igazából arra semmilyen magyarázatot sem adnak, hogy miért éppen ezt tartják az értelmes tervező keze munkájának. Honnan tudjuk, hogy éppen az volt az "eredeti", nem egy másik? Miért nem éppen a Bdellovibrio bacteriovorus ostorát tervezte az értelmes tervező? Hogyan jött létre a többi? Mert ugye ha evolúciós folyamatok során, mutációkkal-szelekcióval, akkor tulajdonképpen az "egyszerűsíthetetlenül összetett" baktérium ostor egy adott alkatrészének a nyolctizedét evolúciós folyamatok alakították ki. Miből gondoljuk, hogy a többit nem? Ha az egyes alkatrészek ilyen mérvű változásokat képesek elviselni és a rendszer mégis működőképes marad, miért tűnik olyan lehetetlennek, hogy eleve ezek a változások alakították ki? Ha pedig feltételezzük, hogy az értelmes tervező tervezte ezt a rengeteg különféle ostort, minek fáradt ennyit? Nekem valahogyan túl sok fölösleges munkának tűnik FlgD fehérjék ezreit, tízezreit megtervezgetni, de ezzel az ÉRTEM hívei is így vannak. Ha emlékszünk még: "Egy intelligens cselekvő számára logikus és egyszersmind hatékony is az élőlényeket közös biokémiai alapon megtervezni." (http://ertem.hu/content/view/173/34/) Viszont akkor miért nem így cselekedett a feltételezett értelmes tervező?
Nyilván nem tudjuk pontosan, minden kétséget kizáróan bizonyíthatóan megmondani, hogyan alakult ki a baktérium ostora, mert olyan élőlényekről van szó, akikből kövületek nem maradnak fenn, ráadásul a vizsgálni kívánt folyamat valamikor három- négymilliárd éve történt, az akkori köztes alakok nyilván azóta rég kihaltak. Akármilyen részletes modellt vázolnánk is fel, meg lehetne kérdezni, hogy jó, jó, szép, szép, akár így is történhetett volna, de mi bizonyítja, hogy tényleg így történt? A válasz nyilván az, hogy ezt sajnos nem tudjuk bizonyítani, ez csak egy lehetséges modell lenne. Viszont ha feltételezzük, hogy evolúciós folyamatok alakították ki, abból következik hogy kellettek lenni egyszerűbb felépítésű ostoroknak is, Behe professzor elmélete pedig bevallottan arra épül, hogy ilyenek nem létezhettek. Viszont mivel ilyen egyszerűbb ostorokat sikerült találni, Behe professzor elmélete hibás, az elméletből következő előrejelzéseket maga a valóság cáfolja.
A fenti gyors körképből egyértelműen kitűnik, hogy "a baktérium ostor" nem egyszerűsíthetetlenül összetett rendszer. Ha megvizsgáljuk Behe professzor többi példáját, ugyanezt tapasztaljuk:
A véralvadási kaszkád fehérjéi is az egyszerűsíthetetlenül összetett rendszerek közé tartoznak Behe professzor szerint, ám 1998 –ban, két évvel a Darwin fekete doboza megjelenése után kiderült, hogy a bálnákból hiányzik a XII –es véralvadási faktor. A génje megtalálható, ám két mutáció működésképtelené tette, pszeudogén (Semba U. és munkatársai 1998.). 2003 –ban pedig kiderült, hogy a gömbhal (Fugu rubripes) véralvadási kaszkádjából öt fehérje hiányzik, amelyek az emlősökben megvannak (Jiang Y és munkatársai 2003.).
Az adaptív immunitást is ide sorolta Behe professzor az eredeti könyvében, ám azóta többen is tanulmányozták az állkapocs nélküli gerincesek immunrendszerét, és meglepő különbségeket fedeztek fel: Ezekben az élőlényekben is véletlenszerű szomatikus rekombinációval alakulnak ki az immunsejtek receptorai, ám különbözik a felhasznált szignálszekvencia, nyoma sincs a magát a rekombinációt katalizáló az állkapcsos gerincesekben meglévő RAG1 és RAG2 enzimeknek, valamint a receptorok teljesen különböző szerkezetűek, az állkapcsos gerincesek immunglobulin doméneket tartalmazó receptora helyett az állkapocsnélküli gerincesekben leucin- gazdag repeat doméneket tartalmaznak a szomatikus rekombinációval előálló receptorok. (Guo P. és munkatársai 2009.)
Az eukarióták csillói is Behe eredeti példái között voltak, sajnos ezekből is ismert egyszerűbb változat. (Woolley D. M. 1997.)
Az elektronszállításról már írtam, ez sem egyszerűsíthetetlenül összetett.
A telomérák fenntartása is egyszerűsíthetetlenül összetett rendszer Behe professzor szerint, nagy kár, hogy az ecetmuslicában például nem az emlősökben megismert telomeráz enzimen alapuló rendszer tartja fenn a kromoszómák végein található szakaszokat, hanem egy módosult, megszelídített ugráló genetikai elem. (Capkova Frydrychova R. és munkatársai 2008.)
Lényeg, ami lényeg, ha egy egyszerűsíthetetlenül összetett rendszerből létezik egyszerűbb változat, az önmagában agyoncsapja a rendszer egyszerűsíthetetlenségét és bizony az erre felhozott példák mindegyikéről kiderült időközben, hogy bizony léteznek egyszerűbb változataik is, azaz nem egyszerűsíthetetlenül összetettek. Hogy újra idézzem az ÉRTEM hivatalos honlapját: "Egyszerűsíthetetlen összetettség nem állhat elő fokozatosan, egy korábbi rendszer jelentéktelen, egymás utáni változásainak eredményeként, mert definíció szerint egy egyszerűsíthetetlenül összetett rendszernek bármely őse működésképtelen." Erre mondják, hogy ipi-apacs. Az úgynevezett "egyszerűsíthetetlenül összetett" rendszerek nem azok és kész.
Egyetlen kérdést nem érintettünk csak eddig, azt a tényt, amelyet sehol sem fektettek le írásban. Annak idején leveleztem az ÉRTEM hazai apostolaival, akkor döbbentem rá erre a kimondatlan törvényre. Ha bárki más rendszert próbál példaként állítani, mint amelyeket Behe professzor említett annak idején a könyvében, rögtön megmagyarázták, hogy ez csak egy szokásos evolucionista csalás, egy nyilvánvalóan nem-egyszerűsíthetetlenül összetett rendszert veszünk és arról bizonyítjuk, hogy nem is egyszerűsíthetetlenül összetett. Ekkor jöttem rá, hogy az egyszerűsíthetetlenül összetettségnek létezik egy mindennél fontosabb követelménye is, amelyet azonban sohasem foglaltak írásba: Az adott rendszert az ÉRTEM apostolai ismerjék el annak.
Mondanék egy nagyon egyszerű példát, az ember szaporodását. Ugye mi méhlepényes emlősök vagyunk, az utódaink az anya méhében fejlődnek kilenc hónapig, csak ez után születnek meg, az újszülötteket ez után még hosszú ideig az anya tejével táplálja. Ez a rendszer, vagyis az ember szaporodása az egyszerűsíthetetlenül összetett rendszer minden követelményét kielégíti: Egységes rendszer, számos egymással kölcsönható elemből áll (petefészkek, méh, méhlepény, tejmirigyek, az egész folyamatot szabályozó hormonok, ezek receptorfehérjéi), amelyek mindegyike hozzájárul az alapvető működéshez és amelyek közül bármelyik eltávolítása működésképtelenné teszi az egész rendszert. Ezen kívül számos előnye van még ennek a példának: Gyakorlatilag mindenki ismeri valamennyire a működését, hiszen egy bizonyos koron túl már mindenki tisztában van azzal, hogyan is lesznek a kisbabák. Eléggé jól kutatott rendszer, számos emlősállat szaporodását ismerjük részletesen, nyilván számos példát lehetne rá hozni. Ráadásul itt a rendszer minden egyes elődjének működőképesnek kell lennie, hiszen egy szaporodásképtelen állat utódok híján nyilvánvalóan nem lehet a mai emlősök őse.
Az evolúció elmélete feltételezi, hogy ez a bonyolult, minden jel szerint egyszerűsíthetetlenül összetett rendszer a hüllők szaporodásából fejlődik ki, akik csak raknak egy tojást és kész. Ugyanúgy elő lehetne hozni az érveket, miszerint az ember szaporító szervrendszerének nem lehettek működőképes elődei, hiszen bármelyik elemének eltávolítása az egész rendszert működésképtelenné teszi és ugyanígy fel lehetne hozni az összes szokásos érvet, majd levonni a következtetést, hogy az értelmes tervező tervezte. Igazán tetszetős lenne, nem? Mégiscsak sokkal de sokkal közérthetőbb lenne, mint a baktérium ostorral példálózni, amiről lássuk be, a legtöbben azt sem tudják eszik –e vagy isszák?
Azonban van egy apró bökkenő, nevezetesen az emlősök ősei nem haltak ki mind, a hüllők szaporító szervrendszere és az emlősök szaporító szervrendszere közti átmeneti alakok közül ma is sok él. A tojásrakó emlősök lágy héjú tojásokkal szaporodnak, de a kikelő utódaikat tejmirigyeik váladékával táplálják. Az erszényes emlősök utódai méhlepény hiányában csak rövid ideig fejlődhetnek az anyaméhben, így igen fejletlen állapotban kell megszületniük, az anyaállat erszényében fejlődnek aztán önállóan is életképes utódokká. Vagyis egy ilyen bonyolult egyszerűsíthetetlenül összetett rendszer is kialakulhat lépésenként, apróbb változtatásokkal, egyszerűbb ősökből. Akkor tulajdonképpen miért gondoljuk, hogy a többi nem? Ugye lehet azzal érvelni, hogy az ember szaporítószervrendszere nem egyszerűsíthetetlenül összetett, ez csak egy hajánál fogva előrángatott példa, amit szándékosan hoztam. Viszont akkor jó volna megmagyarázni, hogy miért nem az? Van esetleg ezeknek a meghatározásoknak egy titkos záradéka, amit kívülállók nem ismerhetnek meg, csak Behe professzor? Erre a rendszerre az eredeti és a bővített meghatározások mindegyike ráillik, egyetlen "bűne" van, hogy tudjuk, hogyan alakult ki, mivel az ősi, egyszerűbb rendszerek közül több fennmaradt a mai napig. Ha teszem azt már kihaltak volna a tojásrakó emlősök és az erszényesek, vajon ez is egyszerűsíthetetlenül összetett rendszer lenne? A személyes véleményem az, hogy az a bizonyos titkos záradék egyszerűen annyi, hogy csak az a rendszer egyszerűsíthetetlenül összetett, amelyről nem tudjuk pontosan, hogyan alakult ki. Behe eredeti meghatározása számos rendszerre tökéletesen illik, amelyek kialakulását ismerjük, ezek azonban nem számítanak valódi egyszerűsíthetetlenül összetett rendszernek, hiszen tudjuk, hogyan alakultak ki. Nem mellesleg ez a körkörös érvelés szép példája: Nem tudjuk hogyan alakultak ki az egyszerűsíthetetlenül összetett rendszerek, hiszen csak azt ismerjük el annak, amiről nem tudni, hogy alakult ki, aminek a kialakulása ismert, azok nem egyszerűsíthetetlenül összetettek. Amennyiben nincs igazam, felkérném a kritikusokat, hogy magyarázzák meg, miért nem egyszerűsíthetetlenül összetett az ember szaporítószervrendszere, vagy bármely más olyan rendszer, amelyre tökéletesen ráillik Behe professzor meghatározása, de mégsem fogadják el annak! "Az egyszerűsíthetetlen összetettségre valójában egy biokémikus tankönyv valamennyi oldalán találhatunk példát." (http://ertem.hu/content/view/31/16/)
Összefoglalva Behe professzor sokszor varázsszóként emlegetett "egyszerűsíthetetlenül összetett" rendszerei tulajdonképpen a rések istene ("god of the gaps") érvelés újabb köntösben. A tudásunknak mindig lesznek házagai, ha úgy tetszik határai. Mindig lesznek folyamatok, amelyeket nem ismerünk teljes részletességgel, vagy egyáltalán nem, mert eddig még nem kutatta senki. Kijelenthetjük, hogy ott lakik Isten, de annál kínosabb, ha a tudásunk határai kicsit kijjebb kerülnek, például új felfedezések nyomán és kiderül, hogy mégsem. Behe professzor minden példaként hozott egyszerűsíthetetlenül összetett rendszeréből ismerünk egyszerűbb, ám működő változatokat, az elmélete egyszerűen megdőlt. Ezek után meglepő, hogy mégis tizenhárom éve töretlen lendülettel véres kardként hordozzák körbe az ÉRTEM hívei, mint a hitük fő bizonyítékát, annak ellenére, hogy az elméletet gyakorlatilag megjelenésének pillanatában meg is cáfolták biológusok, ráadásul az azóta napvilágra került tények újra és újra csak Behe professzor elméletét cáfolják.
Capkova Frydrychova R, Biessmann H, Mason J M. (2008): Regulation of telomere length in Drosophila; Cytogenetic and Genome Research 122(3-4):356-64
Guo P, Hirano M, Herrin BR, Li J, Yu C, Sadlonova A, Cooper MD (2009): Dual nature of the adaptive immune system in lampreys; Nature 459(7248):796-801
Hizukuri Y., Yakushi T., Kawagishi I., Homma M. (2006): Role of the intramolecular disulfide bond in FlgI, the flagellar P-ring component of Escherichia coli; Journal of Bacteriology 188(12):4190-7
Jiang Y, Doolittle RF. (2003): The evolution of vertebrate blood coagulation as viewed from a comparison of puffer fish and sea squirt genomes; Proc Natl Acad Sci U S A. 100(13):7527-32.
Pallen M. J., Matzke N. J. (2006): From The Origin of Species to the origin of bacterial flagella; Nature Reviews Microbiology 4, 784-790
Semba U, Shibuya Y, Okabe H, Yamamoto T. (1998): Whale Hageman factor (factor XII): prevented production due to pseudogene conversion; Thrombosis Research 90(1):31-7.
Woolley D. M. (1997): Studies on the eel sperm flagellum. I. The structure of the inner dynein arm complex; Journal of Cell Science. 110 ( Pt 1):85-94
Ha az ÉRTEM oldaláról érkeztél ide, kérlek olvasd el a cikksorozat többi részét is!
Nem értik 1. - A nejlonevő baktérium
Nem értik 2. - A polip szeme
Értelmes tervezés - A P-elem
Nem értik 3. - Genetika és makroevolúció
A mutációkról
A fajok eredete
Biokémiai hasonlóságok
Nem értik 5. - A specifikus összetettség
Nem értik 6. - Az értelmes tervezés mozgalom tudományos tevékenysége
 (Vendégmunkásunk, SexComb, újra lesújt.)
(Vendégmunkásunk, SexComb, újra lesújt.)
Cikksorozatunk harmadik részéhez érkezett (előzmények itt és itt), szerencsére az értelmes tervezés hívei ellátnak bőven elemeznivalóval. A ma boncasztalra fektetett cikk itt található, a jelöletlen idézetek ebből az írásból származnak.
Mendelt nem véletlenül tartjuk
a genetika atyjának. Az őáltala végzett kísérleteket a mai napig
tanítják az egyetemeken, éppen azért, mert tiszta kísérleti rendszert
hozott létre, adatokat gyűjtött, majd az eredményeket megfelelően
értékelte ki. Mit is csinált ez a szerzetes? Először is megfelelő
modellszervezetet keresett, amelynek végül is a borsó, azaz a
Pisum sativum bizonyult. A borsó ugyanis képes önmegtermékenyítésre,
gyorsan nő, kevés helyet igényel, ráadásul, mivel termesztett növény,
rengeteg változata ismert. Mendel harminckét borsótörzsből két
év alatt választotta ki azt a néhány vonalat, amellyel kísérletezni
kezdett. Ezek után hét tulajdonságot választott ki, a további munkája
során ezek öröklődését vizsgálta.
A kiválaszott tulajdonságok:
Ezek után az egyes törzseket addig keresztezte, amíg az utódok több nemzedéken át, nagy mintaszám esetén is egyformák voltak, azaz már nem hasadtak (szegregáltak) tovább. Ő ezeket tiszta vonalaknak nevezte, a mai szóhasználattal homozigótáknak hívjuk őket. Egy diploid élőlény ugyanis minden kromoszómájából két példánnyal rendelkezik, az egyiket az egyik, a másikat a másik szülőjétől kapja, így minden egyes génünkből is két példánnyal rendelkezünk. Egy gén különböző változatait alléloknak nevezzük. Mendel a kísérleteiben minden egyes gén két-két allélját vizsgálta. Az általa tiszta vonalaknak nevezett homozigóta növények tehát az adott génnek az egyik allélját örökölték mindkét szülőjüktől, így utódaiknak is csak ezt adhatták át, így a tiszta vonal egyedein belül keresztezve nem hoztak létre más fenotípusú utódokat, azaz az utódnemzedékek nem hasadtak, azaz szegregáltak.
Mendel a kísérleteiben pontosan nyomon követte az egyes egyedek leszármazását, amit úgy végzett, hogy az éretlen borsóvirágokból eltávolította a porzókat, majd egy kis papírtölcsérrel lefedte őket, hogy idegen virágpor ne szennyezhesse a kísérletét. A kísérletében használt tiszta vonalakat, azaz homozigótákat nevezte P, azaz szülői (Parentes) nemzedéknek, míg az ő utódaikat F1, az F1 nemzedék egymás közötti keresztezéséből származó utódait F2 nemzedéknek nevezte el.
Ezek után tiszta vonalú (!)
piros virágú borsót keresztezett tiszta vonalú fehér virágú borsóval
(P – szülői nemzedék). Azt tapasztalta, hogy az utódaik, az F1
nemzedék egyöntetűen piros virágúak lettek. Erre magyarázatot
is adott, a két szülő a virágszínt meghatározó génnek különböző
allélját hordozta, ám mindkettő ugyanabból az allélból kettőt.
Az utódaik mindkét különböző allélből egyet – egyet kaptak,
így egyformák lettek, mivel a piros virágszín domináns a fehér
felett, így a növények fenotípusát ez határozza meg. Gregor Mendel
jelölésével élve: A tiszta vonalú, piros virágú szülő AA, míg
a tiszta vonalú, fehér virágú szülő aa allélokat hordozott. Az
F1 utódnemzedék egyedei az egyik szülőtől egy A allélt kaptak,
mivel nem kaphattak mást, míg a másik szülőtől egy a allélt kaptak,
hiszen tőle sem örökölhettek mást. Így az F1 nemzedék genotípusa
Aa lett. Ez alapján könnyű meghatározni azt is, hogy ugyanannak
a génnek két allélja közül melyik a domináns és melyik a recesszív: domináns az, amely a heterozigóták fenotípusát meghatározza, recesszív
az, amely fenotípusa csak homozigóta formában jelentkezik.
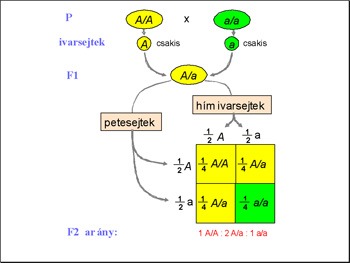 Ezek után az F1 nemzedék
egyedeit egymással keresztezte és meglepődve tapasztalta, hogy a
körükben ismét megjelentek fehér virágú egyedek is. Mendel lángelméje
ott mutatkozott meg, hogy meg is számolta az utódokat, azaz statisztikai
alapon elemezte őket! 705 darab piros virágú és 224 darab fehér
virágú növényt kapott. Ezt az arányt ő 3:1 –nek mérte. Ezek
után kíváncsi volt rá, hogy az utódok vajon tiszta vonalúak –e?
Az F2 nemzedék egyedeit önmegtermékenyítéssel szaporította és
megvizsgálta az utódaikat (F3 nemzedék). Azt tapasztalta, hogy a
domináns fenotípust mutató egyedek harmada tiszta vonalú, azaz ebben
az esetben az utódaik is piros virágúak lesznek, míg a kétharmaduk
nem tiszta vonalú, azaz az utódaik között fehér virágúak is felbukkannak,
míg a fehér virágúak mind tiszta vonalúak. Arra következtetett,
hogy a piros virágúak egyharmada, azaz az összes utód egynegyede
AA genotípusú, azaz tiszta vonalú, kétharmada, azaz az összes utód
fele Aa genotípusú, azaz fenotípusosan piros virágú, ám az utódaik
között fehér virágúak is megjelennek, míg a fehér virágúak
aa genotípusúak, természetesen tiszta vonalúak.
Ezek után az F1 nemzedék
egyedeit egymással keresztezte és meglepődve tapasztalta, hogy a
körükben ismét megjelentek fehér virágú egyedek is. Mendel lángelméje
ott mutatkozott meg, hogy meg is számolta az utódokat, azaz statisztikai
alapon elemezte őket! 705 darab piros virágú és 224 darab fehér
virágú növényt kapott. Ezt az arányt ő 3:1 –nek mérte. Ezek
után kíváncsi volt rá, hogy az utódok vajon tiszta vonalúak –e?
Az F2 nemzedék egyedeit önmegtermékenyítéssel szaporította és
megvizsgálta az utódaikat (F3 nemzedék). Azt tapasztalta, hogy a
domináns fenotípust mutató egyedek harmada tiszta vonalú, azaz ebben
az esetben az utódaik is piros virágúak lesznek, míg a kétharmaduk
nem tiszta vonalú, azaz az utódaik között fehér virágúak is felbukkannak,
míg a fehér virágúak mind tiszta vonalúak. Arra következtetett,
hogy a piros virágúak egyharmada, azaz az összes utód egynegyede
AA genotípusú, azaz tiszta vonalú, kétharmada, azaz az összes utód
fele Aa genotípusú, azaz fenotípusosan piros virágú, ám az utódaik
között fehér virágúak is megjelennek, míg a fehér virágúak
aa genotípusúak, természetesen tiszta vonalúak.
Ezzel a keresztezéssel Mendel "belelátott" az F2 nemzedék piros virágú egyedeinek a genomjába, meg tudta állapítani, mi volt a genotípusuk. Az utódok megoszlása pontosan követte a modell alapján számolt matematikai esélyeket. A piros virágú F1 nemzedék tagjai mind Aa genotípusúak voltak, azaz az utódaik fele az A, másik fele az a allélt örökölte tőlük. Így a modell alapján megjósolható az F2 nemzedék tagjainak a fenotípusa. (Az ábra innen származik.)
Mivel az F2 nemzedék tagjai egyenlő eséllyel öröklik a szülői alléleket, azaz mindkét szülőjüktől 1/2 eséllyel kapják meg az A vagy az a allélt. Tehát ha az első társaságot nézzük, mi az esélye, hogy AA genotípusú egyed jöjjön létre? 1/2 eséllyel örökli az egyik szülőjétől az A allélt, 1/2 eséllyel örökli a másik szülőjétől is az A allélt. Mivel a mindkét esemény véletlennek tekinthető, az együttes bekövetkezésük valószínűségéhez az egyes valószínűségeket össze kell szorozni. 1/2X1/2=1/4, azaz a matematikai modell alapján az F2 nemzedék egynegyede lesz tiszta vonalú piros virágú, azaz AA genotípusú. Ugyanígy kiszámolható, hogy az F2 nemzedék egyedei közül hányan lesznek fehér virágúak, azaz aa genotípusúak: 1/2X1/2=1/4. Ezek után kiszámolható, hogy 1/2 az esélye az Aa genotípus kialakulásának.
Mendel kísérletei pontosan követték a matematikai modellt, azaz tiszta vonalú szülők (P) AA X aa keresztezésekor az F1 utódok egyöntetűen Aa genotípusúak, fenotípusukra nézve a domináns allél által meghatározott fenotípust mutatják, azaz piros virágúak. Az ő beltenyésztésükből származó utódok 1/4 –e AA azaz homozigóta piros virágú, 1/2 –e Aa azaz heterozigóta piros virágú és 1/4 –e aa genotípusú azaz fehér virágú. Azaz az F2 nemzedék fenotípusára nézve 3/4 –ük piros (AA+Aa), 1/4 –ük fehér virágú (aa).
Ezek után megismételte a kísérletet az általa vizsgált tulajdonságokkal és mindegyikkel hasonló eredményt kapott. Ebből arra következtetett, hogy az általa feltárt törvényszerűségek nem egyedi, elszigetelt példák, hanem a növények öröklődésére általánosan jellemzők. Azóta beigazolódott, hogy az általa feltárt folyamatok nem csak a növények öröklődésére igazak, hanem minden diploid szervezetére.
| Szülői fenotípusok | F1 | F2 | F2 arány |
| Piros x fehér virág | mind piros | 705 piros : 224 fehér | 3,15 : 1 |
| Sárga x zöld szem | mind sárga | 6022 sárga : 2001 zöld | 3,01 : 1 |
| Felfújt x befűzött hüvely | mind felfújt | 882 felfújt : 299 befűzött | 2,95 : 1 |
| Zöld x sárga hüvely | mind zöld | 428 zöld : 152 sárga | 2,82 : 1 |
| Axiális x terminális virág | mind axiális | 651 axiális : 207 terminális | 3,14 : 1 |
| Hosszú x rövid szár | mind hosszú | 787 hosszú : 277 rövid | 2,84 : 1 |
Mendel éppen azt igazolta, hogy az egyes allélok öröklődése matematikai esélyeket követ, a recesszív allélok nem lappanganak valamilyen varázslatos módon néhány száz nemzedéken át, hogy aztán látszólag a semmiből újra előtörjenek. Az allélek "újrakeveredése" szigorú statisztikai törvényszerűségeket követ. Két heterozigóta keresztezésekor az utódok negyede a recesszív fenotípust mutatja. Ha egy adott fenotípusért két függetlenül öröklődő gén recesszív allélja a felelős, az utódok 1/16 –oda akkor is a recesszív fenotípust mutatja (1/4X1/4), ha három gén recesszív allélja felelős érte, akkor 1/64 –ede (1/4X1/4X1/4) és így tovább. Az egyes egyedek genotípusa egyszerű keresztezési kísérletekkel vizsgálható, így a "rejtett" recesszív allélek megléte egyértelműen igazolható vagy cáfolható. Mendel éppen azt igazolta, hogy megfelelő kísérleti rendszerben az egyes egyedek genotípusa éppen úgy vizsgálható, mint a fenotípusa.
Vizsgáljuk meg Gregor Mendel
eredményeinek fényében az értelmes tervezés híveinek állításait:
"A legtöbb variáció
a meglévő gének rekombinációjával (újra-elrendeződésével)
keletkezik."
"Az
„új” jelleg, ami látszólag a semmiből bukkan elő, valószínűleg
nem is új, csupán egy recesszív gén kifejeződése, amely mindvégig
jelen volt. Amikor a tenyésztő újfajta kutyát vagy húsosabb marhát
hoz létre, valójában csak a géneket keveri meg egy kicsit, hogy
a recesszív gént kifejezésre juttassa."
Alighanem Mendel eredményeinek
a fényében el kell, hogy vessük azt a feltételezést, hogy évszázadokon
át, több százezer egyed genomjában százszámra lappanghatnának
fel nem ismert recesszív allélok, anélkül, hogy egyetlen homozigóta
recesszív egyed is született volna. Egy allél hiába recesszív,
az utódoknak átadása szigorú matematikai törvényszerűségeket
követ, ezek mérhetőek, százalékos esélyek formájában kifejezhetőek,
ezek alapján az egyes recesszív jellegek fenotípusbeli megjelenése
pontosan megjósolható. Hol vannak a tacskólábú farkasok? Hol vannak
a bulldogpofájú farkasok? Miért nem születnek pincsiméretű farkasok?
Ha ezek az allélek olyan gyakoriak, hogy a tenyésztők "vakon",
azaz a domináns fenotípust mutató egyedek tenyésztésével is mintegy
"véletlenül" begyűjtötték a tenyésztett kutyák közé,
a vadonban miért nem figyelhetőek meg a recesszív allélekre homozigóta
egyedek? Rengeteg farkast tartanak állatkertekben, mégsem születnek
köztük a szerző által feltételezett recesszív jellegeket hordozó
egyedek, pedig, ha ezek a gének ott lappanganának a genomjukban, időnként
mindenképpen kellene születnie egy-egy recesszív fenotípust is mutató
egyednek. Viszont legjobb tudomásom szerint nem írták le a farkasokról,
hogy ilyen elképesztő formagazdagságot mutatnának, márpedig, ha
ezek a recesszív allélek ott lennének a farkas genomban, éppen a
mendeli genetika szabályai szerint kiszámítható eséllyel születniük
kellene a recesszív fenotípusokat mutató egyedeknek. Viszont valamiért
mégis hiányoznak a recesszív allélekre homozigóta egyedek. Hogyan
lehetséges ez? A legvalószínűbb magyarázat annyi, hogy a tenyésztés
során bizony új mutációk is keletkeztek, nem csak a már meglévő
allélek keveredtek újra.
Amúgy érdekes kérdés, hogy
a feltételezett értelmes tervező minek tervezett ennyi fölösleges,
sőt, káros allélt az egyes génekre, ha amúgy sem nyilvánulnak
meg soha a vadállatok fenotípusában? Feltételezem az értelmes tervezés
nem azért történt, hogy az ebtenyésztők egyesületi tagjainak legyen
megfelelő alapanyaga majdan, százezer évekkel később.
"Vegyük szemügyre
a zsiráfot: túlméretezett végtagok, megnyúlt nyak, esetlen testtartás
– látszólag minden ügyetlenül aránytalan. Ennek ellenére a zsiráf
testrészei bámulatos összhangban
állnak egymással: az állat elegáns könnyedséggel mozog,
és olyan erős a rúgása, hogy kevés természetes ellensége van."
A zsiráf nagy méretű, vadon
élő, lassan szaporodó afrikai állat. Szinte mindenki ismeri, viszont
tökéletesen alkalmatlan genetikai modellszervezetnek. Több mint egy
évig vemhes, a nőstény három-négyévesen válik ivaréretté, azaz
egyetlen nemzedékváltást kb. öt év alatt lehetne megfigyelni, ráadásul
rendszerint egyetlen egyet borjadzik, így az utódok statisztikai elemzése
is elég nehézkes. Ennek fényében nem meglepő, hogy tudomásom szerint
még soha senki sem végzett egyetlen genetikai kísérletet sem zsiráfon.
Vitaindítónak viszont tökéletes egy alapvetően genetikai tárgyú
cikkben, hiszen mivel egyetlen sornyi adat sincs az öröklődéséről,
a szabad elméletalkotást nem zavarják holmi tények és kísérleti
eredmények. (Néhány elszórt vizsgálat történt, mikroszatellita
merkereket, mitokondriális DNS-t vizsgáltak, illetve egy interferon
gént, azonban a zsiráf öröklődéséről nincsenek adatok.)
 Nézzünk meg inkább egy példát,
ahol a háziasítás folyamata jól követhető sőt, még az egyes
tulajdonságok öröklődése is ismert. Gondolom nem kell bemutatnom
a szíriai aranyhörcsögöt, amely kedvelt háziállat nálunk is.
Latin neve Mesocricetus auratus. Magyar nyelven is bőséges
forrásanyag áll a rendelkezésünkre róla (pl. itt,
itt, itt, vagy
itt), így nem fáradnék
a bemutatásával.
Nézzünk meg inkább egy példát,
ahol a háziasítás folyamata jól követhető sőt, még az egyes
tulajdonságok öröklődése is ismert. Gondolom nem kell bemutatnom
a szíriai aranyhörcsögöt, amely kedvelt háziállat nálunk is.
Latin neve Mesocricetus auratus. Magyar nyelven is bőséges
forrásanyag áll a rendelkezésünkre róla (pl. itt,
itt, itt, vagy
itt), így nem fáradnék
a bemutatásával.
Miért ilyen különleges ez a rágcsáló? Az első aranyhörcsögöt 1930–ban fogták be, ez egy anyaállat volt az éppen meglévő almával együtt. A fiatalok közül mindössze három egyedet, egy nőstényt és két hímet tenyésztettek tovább, a ma fogságban élő aranyhörcsögök ettől a három egyedtől származnak (Később alapítottak néhány más tenyészállományt is, újabb befogott egyedekből, de ezekkel a populációkkal nem foglalkoznék.). A vad típusú hörcsög – meglepő módon - aranyszínű bundájú, ám az idők során a tenyésztők több különböző színváltozatot alakítottak ki. Ezek közül csak néhányat tárgyalnék, természetesen a teljesség igénye nélkül.
Létezik egy halványszürke színváltozat, amelyet az lg gén domináns allélja (Lg) okoz. Ismert egy ezüstszürke változat, amelyet az sg gén domináns allélja (Sg) okoz. A sárga színt az X kromoszómához kötött to gén domináns alléja (To) alakítja ki. Ezeken kívül léteznek a bunda mintázatát módosító gének is. A ragyogó szatén színt az sa gén domináns allélja (Sa) okozza. Ezeken kívül domináns jelleg a háton végigfutó fehér csík (ba gén Ba allélja), illetve a bunda fehér foltossága (ds gén Ds allélja), valamint a fehér has is (wh gén Wh allélja). Mivel az összes szelídített aranyhörcsög attól a nyolcvan éve élt három jószágtól származik, kijelenthetjük, hogy ezek az allélok bizony mind a szelídítés során jelentek meg, új mutációs események nyomán, egyszerűen azért, mert dominánsan öröklődnek, azaz az alapító egyedek nem hordozhatták őket a genomjukban észrevétlenül, mindenképpen megjelent volna a fenotípusukban is, azaz látni kellett volna, ha ezeket az allélokat hordozzák. Az aranyhörcsög esetében kizárhatjuk, hogy vad populációkkal történt volna valamilyen keveredés, hiszen fogságban tenyésztik, ráadásul vadon csak a világ egy szűk területén él. Ezzel a rövidke példával azt hiszem a boncolgatott cikk minden kiindulási állítását sikerült cáfolnom, egy megfelelő modellállat segítségével:
"Ezzel szemben a mendeli
modellben a gének sokkal inkább különálló
részecskékhez hasonlítanak, amelyek az
öröklődés folyamán nem változnak."
" Elfogadható becslés, ha azt mondjuk, hogy egy 1000 fős populációban
egyetlen egyedben egyetlen
mutáció fordul elő."
"Hogyan megy végbe
a változás a mendeli genetika keretein belül?"
Drake J. W. és munkatársai
1998–as cikkében találunk néhány érdekes adatot az egyes szervezetekben
megfigyelt spontán, azaz minden külső beavatkozás nélkül bekövetkező
mutációk gyakoriságáról. A spontán mutációk bekövetkezésének
esélye egy szervezeten belül az egyes gének esetében sem egyforma,
például kukoricában a spontán mutációk bekövetkezésének esélye
a több gén átlagában: 7,7X10-5 . Ennek fényében az
1X10-3 becslés, amit a szerző hoz éppen csak százszor-ezerszer
több, mint a valóságban bekövetkező mutációk száma. Ezzel meg
is fejtettük az okát, miért bizonyultak olyan állandónak a gének
Mendel kezeiben: Ő a kísérleteiben néhány száz növényt vizsgált
végig, a legnagyobb egyedszámot is a borsószemek vizsgálatakor elemezte,
körülbelül nyolcezer egyedet. Kukoricában átlagosan körülbelül
minden hétszázhetvenezredik egyedben történik mutáció egy adott
génben, tehát matematikai esélye akkor lett volna, hogy Mendel egyetlen
egy mutációs eseményt megfigyeljen a borsószem színében, ha körülbelül
százszor ennyi egyedet vizsgált volna a kísérletben, a többi tulajdonság
vizsgálatakor pedig még ennél is kisebb mintaszámmal dolgozott.
Ezek alapján elég könnyű megfejteni ezt a rejtélyt: Spontán mutációk
lényegesen ritkábban történnek, mintsem hogy Mendelnek esélye lett
volna meglátni őket a néhány száz egyeddel végzett kísérleteiben,
így lehet, hogy ő a géneket megváltoztathatatlan egységnek látta.
Egyszerűen túl kevés egyedet vizsgált ahhoz, hogy mutációs eseményeket
figyelhessen meg. Ugyanezen cikk szerint egy egérben 6,6X10-6
ecetmuslicában 8X10-6 eséllyel történnek mutációk.
Ez alapján akárhogyan is nézzük, a szerző becslése lényegesen
több mutációval számol, mint amennyi a valóságban megtörténik.
" Továbbá
a beltenyésztés a hibás jellegek koncentrációját eredményezheti,
és ahogy a faj átlagos morfológiájától egyre
távolabb kerülünk, egyre több disszonanciát, stresszt
és csökkenő termékenységet tapasztalunk.
Az ilyen túltenyésztett populáció
gyakran hajlamos visszafejlődni a faj
átlagos morfológiai irányába."
Ha a szerző maga szögezi
le, hogy új allélok nem keletkeznek, csak a meglévőek eloszlása
változik, hogyan fejlődhetnének vissza a beltenyésztett populációk?
A szerző – nem mellesleg teljesen alaptalan - állítása szerint
minden egyes tenyésztett állatban meglévő minden egyes jelleg recesszíven
öröklődik, a tenyésztők mindössze ezeket a recesszív alléleket
hordozó egyedekből hoznak létre tiszta vonalakat, azaz állítják
homozigóta állapotba. Amennyiben az adott, túltenyésztett jelleget
okozó tulajdonság recesszíven öröklődik, akkor nem változhat
meg mutációs esemény nélkül, hiszen az egyed homozigóta, azaz
"aa" genotípusú, onnan tudjuk biztosan, mert különben
a domináns fenotípust mutatná. Ebben az esetben teljesen érthetetlen,
hogyan "hajlamos az ilyen túltenyésztett populáció
visszafejlődni a faj átlagos morfológiai irányába", hiszen
ehhez az egyedeknek valahonnan varázslatos módon génenként egy domináns
allélt kéne szerezniük a semmiből, hiszen az ő genomjukban ez nem
lehet meg. Hogyan lehetséges ez? Mert ugye ha ez bekövetkezne, akkor
éppen az a hasznos mutáció lenne, amely az értelmes tervezés hívei
szerint nem is létezik.
" A mutációk létrehozhatnak
például összegyűrődött, túl- vagy
alulméretezett szárnyakat. Eredményezhetnek megduplázódott szárnyakat
is, de nem eredményeznek újfajta szárnyakat,
és főleg nem alakítják át a gyümölcslegyet egy
újfajta rovarrá."
" Nincs azonban az
a mennyiségű finombeállítás, ami a jelenlegi testfelépítéséből
egy teljesen új testfelépítést tudna létrehozni."
Az ilyen kijelentések előtt
érdemes lenne tisztázni, mi számít "újfajta" szárnynak?
Ha az nem, ami nagyobb, az sem ami kisebb, az sem amiből kettő helyett
négy van? Nem tudom, nekem az, hogyha a légynek két szárnya helyett
négy nő, eléggé új testfelépítés. Például ha egy ember négy
karral születne, vagy a kézfejei közvetlenül a vállából állnának
ki, esetleg a karjai leérnének a földig, vagy a karjai helyett is
lábai nőnének, nem mondanám rá, hogy ez teljesen olyan, mint a
többi, semmi új nincs benne. Sokkal de sokkal könnyebb lenne érdemi
vitát folytatni, ha az értelmes tervezés hívei meghatároznák az
elvárásaikat. Ha eldöntenék, hogy mi számít "újfajtának",
akkor lehetne rá példát hozni. A jelenlegi helyzet az, hogy az "újfajta"
meghatározása annyi: Amilyen mutációt valaha is leírtak, az éppen
nem "újfajta". Mikor lenne elfogadható az eredmény, azaz
mik az "újfajta" követelményei? Sajnos erre semmilyen elfogadható
meghatározást sem kapunk, ami, ha meggondoljuk valóban kényelmes,
hiszen így akármilyen példát hozna is egy biológus, arra rá lehet
mondani, hogy pont nem az.
"Másképpen fogalmazva,
a tenyésztők édesebb kukoricát, húsosabb tehenet tudnak produkálni,
de nem tudják a kukoricát más növénnyé
vagy a marhát más állattá alakítani."
Érdemes megvizsgálni a Raphanobrassica
nevű növényt. Ezt
Gregorij Karpecsenkó szovjet tudós hozta létre 1928–ban, tehát
nem igazán új eredmény. Retek és káposzta növényeket keresztezett,
ám úgy, hogy mindkettőt kolhicinnel kezelte, így a kromoszómák
nem váltak szét az ivarsejtképzéskor és az utódba mindkét növény
teljes genomja bekerült. A retek és a káposzta diploid kromoszómaszáma
egyaránt 18, az új növény diploid sejtjeiben 36 kromoszóma található.
Amiért érdekes: Ez a növény egyáltalán nem terméketlen, ivarsejteket
képez, magot hoz, így minden további nélkül termeszthető, azonban
egyik szülői fajjal sem képes termékeny utódot létrehozni. Sajnos
a növény gyökere a káposztáéra, levele pedig a retekére hasonlít,
így nem igazán ehető, de kétségkívül ez egy olyan eset, amikor
a tenyésztők két fajt egy teljesen más fajjá alakítottak, amelyik
külső-belső tulajdonságaiban különbözik mindkét szülői fajtól.
Másik, később végzett növénynemesítési munka a Triticale, azaz
a búza és a rozs teljes genomját hordozó gabona megalkotása, az
ötvenes években. A termesztett búza diploid kromoszómaszáma 42,
a rozsé 14, nem csoda, hogy a Triticale diploid kromoszómaszáma 56.
A Triticale mindkét szülői faj jellegzetességeit magán viseli.
A tenyésztők igenis képesek egyes növényeket "más növénnyé"
alakítani.
"Ezzel szemben a mendeli
modellben a gének sokkal inkább különálló
részecskékhez hasonlítanak, amelyek az
öröklődés folyamán nem változnak."
"A mutációk szerencsére eléggé ritkák, mert túlnyomó többségük ártalmas, és csak néhány
semleges."
"Ugyanígy a
mutációk is csak elvétve javítják a DNS
üzenet minőségét, és túl sűrű
előfordulásuk akár halálos is lehet a szervezetre nézve."
"A gének rekombinációjával
ill. mutációkkal generált kicsiny változások elősegíthetik ugyan
a természetes kiválasztódás által a szervezet
„finombeállítását”, lehetővé
téve, hogy ökológiai környezetéhez
jobban alkalmazkodjék."
"Ezek a behatárolt
változások nem akkumulálódnak, ahogy a darwini evolúciós teória
megkívánná az új makrováltozások létrehozásához."
Ha a cikk elején azt állítja,
hogy Mendel igazolta a gének állandóságát, miért ír folyton mutációkról?
Most akkor mutációk egyáltalán léteznek, vagy nem? Már csak azért,
mert a mutációk létét elég nehéz lenne tagadni. Majd a szerző
maga is elismeri, hogy léteznek előnyös mutációk, amelyek "javítják
a DNS üzenet minőségét", ám néhány sorral feljebb határozottan
állítja, hogy mindegyik mutáció káros, vagy semleges. Akkor most
léteznek előnyös mutációk, vagy nem? Ha a mutációk elősegíthetik
a szervezet "finombeállítását", akkor ezek szerint mégis
vannak előnyös mutációk. Ha állítja, hogy az egyes előnyös mutációk
nem halmozódnak fel, miért beszél éppen a "gének" "finombeállításáról",
így többes számban? Ezek szerint akkor a hasznos mutációk mégis
csak felhalmozódhatnak? Az előnyös mutációk felhalmozódása nem
éppen az az evolúció, ami a szerző szerint nem is létezik? Miért
ennyi az önellentmondás ebben az alig néhány oldalas rövid szövegben?
Mi a legnagyobb bajom ezzel
a cikkel? Az, hogy egyetlen egy adat sincs benne. Ír a zsiráfról,
amely állat genetikáját soha senki sem vizsgálta, valamint a kutyákról,
amelynek öröklődése elég kevéssé tanulmányozott, de egyetlen
adatot sem hoz a kutya génjeiről sem. Egyetlen tulajdonság öröklődéséről
sem ír, csak homályos általánosságokat említ. Ráadásul a cikk
állításai minden alapot nélkülöznek, egyszerűen vonzó közhelyek.
Miért van az, hogy az értelmes tervezés hívei, akik az evolúciós
elméletek elemzésekor a legapróbb részletekre is kísérletes bizonyítékot
követelnek, a saját elméleteik esetében ettől nagyvonalúan eltekintenek?
A szerző teljesen alaptalan tényeket szögez le, majd ezekből von
le messzemenő következtetéseket. Mint bölcseleti munka vagy szépirodalmi
alkotás talán megállja a helyét, de semmilyen kapcsolatban sincs
a valósággal. Olvassuk el, mit is jelent pontosan a tudományos módszertan!
Az éppen vizsgált cikk a
tudományos módszer melyik lépését tartja be? Teljesen alaptalan,
egymásnak részben ellentmondó feltételezésekből és minden kísérleti
bizonyítékot nélkülöző kijelentésekből indul ki. Adatgyűjtést
nem végez. Feltételezéseit valóban kibontja, ám ezek adatok híján
teljesen értékelhetetlenek, csak lógnak a levegőben. Előrejelzéseket
nem tesz, és nem ellenőrzi, hogy az általa felépített feltételezések
milyen viszonyban állnak a valósággal. Ahol az állításai ellenőrizhetőek,
azaz egy megfelelő modellrendszerben, például a szíriai aranyhörcsög
esetében, ellent is mondanak a természetben megfigyelt tényeknek,
azaz az ellenőrzés során elbuknak. Ki kell hogy jelentsük, hogy
tetszetős elmélet, ám a valósághoz semmi köze sincs.
 (Sorozatunk előző
részében azt vettük szemügyre, valóban szükséges-e egy Értelmes
Tervező létét feltételezni ahhoz, hogy a nejlonbontó baktériumok létét
megmagyarázzuk, vagy megteszi-e a jó öreg evolúciós elmélet is. Ez
alkalommal vendégmunkásunk, SexComb, az ÉRTEM egy újabb állításának ered a nyomába, mely
ezúttal a polipok szemében véli egy felsőbb, intelligens terv nyomait
felfedezni.)
(Sorozatunk előző
részében azt vettük szemügyre, valóban szükséges-e egy Értelmes
Tervező létét feltételezni ahhoz, hogy a nejlonbontó baktériumok létét
megmagyarázzuk, vagy megteszi-e a jó öreg evolúciós elmélet is. Ez
alkalommal vendégmunkásunk, SexComb, az ÉRTEM egy újabb állításának ered a nyomába, mely
ezúttal a polipok szemében véli egy felsőbb, intelligens terv nyomait
felfedezni.)
Az állatvilágban rengeteg féle szem található, itt a CriticalBiomass blogon is volt ez már számos alkalommal téma (legutóbb lásd itt). Sőt, az írásom témájául szolgáló, polip szem szerkezetéről is jelent már meg írás errefele, így ezt sem kell nagyon részleteznem, mint ahogy az emberi szemmel való összevetést sem.
De tulajdonképpen miről is szól ez a parázs vita? A kérdés lényege éppen az, hogy bár megejtő a külső és belső hasonlóság, a gerincesek szeme egy inverz szem, azaz a retina fény felőli oldalán idegsejtek találhatóak, amelyek az érzéksejtekben a fény hatására keletkezett ingerület elvezetését végzik, míg alattuk, a fénytől távolabb találhatóak maguk a fényérzékeny sejtek. Ezért nevezik ezt az elrendezést "inverz", azaz fordított szemnek. Ezzel szemben a polip szeme everz szem, itt a fényérzékeny sejtek állnak a beérkező fény felé, alattuk helyezkednek el a fény hatására keletkező ingerületet elvezető idegsejtek. A polip everz szeme és az ember inverz szeme működésében, felépítésében meglehetősen hasonló, ám ez az alapvető szerveződésbeli különbség egyértelműen elkülöníti őket. Ez a kettősség, azaz a szemek működésbeli hasonlósága, de szerkezetbeli eltérése az evolúció elméletével egyszerűen magyarázható, egyszerűen ugyanaz a szelekciós nyomás hatott mindkét élőlénycsoportra, azaz a színeket látó, egy pontra élesíthető szemek nagy előnyt jelentenek, így az ilyen jellegű látószervekkel rendelkező élőlények jobban elszaporodnak. Viszont az egyes élőlénycsoportok közös őseinek a szeme eltérő alapszerkezetű látószerv volt, így ugyanazt a feladatot ellátó szerv egymástól függetlenül két, eltérő alapszerkezetű változatban alakult ki a két eltérő élőlénycsoportban.
Nyilván az everz és az inverz szem felépítése különbözik, nyilván a működésük sem tökéletesen azonos. De nem is ez a kérdés. A kérdés az, hogy az azonos élőhelyen, azonos életmódot folytató mélytengeri polipnak miért van everz szeme és a vele egy környezetben élő mélytengeri halnak miért van inverz szeme (hiszen, bár erről az boncolgatott cikk írója elfeledkezik, az összes gerincesnek az emberéhez hasonló szerkezetű szeme van)? Pontosan ugyanazon az élőhelyen élnek, a tenger mélyén, akaratlagosan egyikük sem emelkedik soha a felszínre, belátható, hogy pontosan ugyanolyan körülmények között, ugyanarra használják a szemüket. Odalent nem kell tartaniuk a túl erős fénytől, ráadásul színek sem igazán látszanak, hiszen gyakorlatilag az egyetlen fényforrás az élőlények által termelt halvány derengés. Ha, mint azt az értelmes tervezés hívei sugallják, a látás minősége az ember és polip szem közti különbség egyetlen oka és magyarázata, miért különbözik mégis az egyaránt mélytengeri környezetben, azonos fényviszonyok közt élő, de különböző rendszertani csoportba tartozó fajok látószerve? A tengerben a felszín közelében, például a korallzátonyokon is élnek egymás mellett polipok és halak, gyakorlatilag nappali fényben. Mégis a felszín közelében élő halaknak is inverz szemük van, míg a felszín közelében élő polipok is everz szemmel rendelkeznek. Itt bizony ragyogó fényben élnek mindketten, színeket is látni. Miért különbözik mégis a látószervük? Ugyanígy feltehetjük a kérdést, hogy a szárazföldön élő csigának miért van everz szeme, amikor a mellette élő béka inverz szemmel rendelkezik? Ugyanolyan környezetben élnek, mégis különböző a szemük. Az evolúció talaján álló magyarázat egyértelmű: a csiga puhatestű, nyilván a szeme is a többi puhatestűére hasonlít, míg a béka gerinces, a szeme a többi gerincesére hasonlít. Ha végignézzük, kiderül, hogy a puhatestűek, bárhol éljenek is, everz szemmel rendelkeznek (már amennyiben van szemük), míg a gerincesek, bárhol éljenek is, inverz szemmel rendelkeznek. Nehéz itt az értelmes tervező kéznyomát felfedezni vagy gondolatmenetét követni.... Miért adott az értelmes tervező minden gerincesnek egyforma alapszerkezetű szemet és miért kaptak a puhatestűek azonos szerkezetű, de a gerincesekétől különböző szemet? Hogy a feltételezett értelmes tervező miért tartja magát a leszármazási kapcsolatokhoz, rejtély.
"A digitális fényképező másféle képet készít, mint amilyet a Szmena tud. Az ilyen típusú és minőségű képek elkészítésének pedig mások a műszaki feltételei, mint a kisgép képeinek. Másféle alkatrészeket is biztosítani kell hozzá, és azokat egy meghatározott módon kell összeszerelni."
Vegyük szemügyre egy másik gerinces szervezet inverz szemét! A varangy látását elég alaposan vizsgálták, az ő szeme az emberétől eltérő képet ad: gyakorlatilag csak a mozgást látja, de azt kevés fénynél is, viszont színlátása gyenge, így az ő látása megfeleltethető a polip látásának, ahhoz lehetne hasonlítani, amit mi a "szemünk sarkából látunk". Mégis inverz szeme van, mint minden gerincesnek. Adja magát a kérdés: miért hasonlít az ember és a béka szemének alapszerkezete, ha a béka látása a polip által látott képhez hasonlít? Talán mégsem azért különbözik a két szem, mert másféle képet készít? Miért tervezett a feltételezett értelmes tervező a digitális fényképezőből is egy lebutított változatot, körülbelül Szmena fényképező minőségűt? Miért használta ehhez a "digitális fényképező" alkatrészeit, ha az ilyen kép készítésének mások a műszaki feltételei és másféle alkatrészek kellenek hozzá?
Érdemes azon is elgondolkodni, hogy azért a békák szeme kicsit másképpen működik, mint az emlősöké. Nevezetesen ők nem úgy élesítik a látásukat egy pontra, hogy a szemlencséjük alakját változtatják meg, hanem a szemlencséjüket mozgatják a szemen belül. Akkor most ez hogy is van, nem csak "egy meghatározott módon" lehet összeszerelni ezt az inverz szemet? Talán ez mégsem egy egyszerűsíthetetlenül összetett rendszer? Mégiscsak van némi tere az evolúciónak?
"A tervezési szemléletet követői azonban meg vannak győződve arról, hogy az élővilág egy értelem működéséről árulkodik, ezért a biológiai szervezetekben található elrendezéseknek ésszerű indokai vannak."
A vizsgált cikkben találunk egy vázlatos ábrát az emberi szem szerkezetéről, amelynek csak egyetlen elemét emelném ki, ez a "látógödör". Bár az ábrán szerepel, a szövegben nem találtam nyomát. Mi is ez a látógödör? Hívják még sárgafoltnak is, vagy ünneplős nevén macula luteának. Ez az a pont az emberi szemben, ahol az éleslátás keletkezik, azaz ha valamire ránézünk, akkor a beérkező fény éppen ide esik, ezt a területet használjuk, ha valamire figyelünk, például jelen pillanatban is a sárgafoltodra vetül e szöveg képe, kedves olvasó. Itt a legsűrűbbek a fényérzékeny sejtek az emberi szemben, az ezen kívül eső területek a mozgás érzékelésében játszanak szerepet, csúnya szóval a perifériás látásban. Miért hívják mégis ezt a területet éppen látógödörnek? Ugyanis itt a keresztmetszeti képen a retina mintegy "elvékonyodik", egy apró bevágás, gödör figyelhető meg benne. Miért is van ez? Ha mikroszkóppal vizsgáljuk ezt a területet, valami ilyesmit látunk (A kép innen származik.)
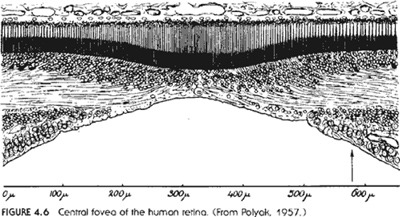 Ezen a
területen ugyanis az idegsejtek mintegy "elhúzódnak"
a fényérzékelő sejtek fölül, így
téve lehetővé az éles látást.
Itt azért felmerül a kérdés, hogy az emberi
szem miért inverz szem, ha azon a ponton, ahol az éles
kép keletkezik, az idegsejtek kénytelenek elhajolni a
fényérzékeny sejtek fölül? Az
idegsejtek léte valóban szükséges a szem
működéséhez, ők továbbítják
a fényérzékeny sejtekben keletkező
ingerületet, de akkor miért éppen arról a
területről húzódnak el (természetesen
nem hiányoznak, a látógödör körüli
idegsejtek nyúlványai továbbítják
az itt található fényérzékeny
sejtek ingerületét), ahol a legjobban kellenének,
hiszen nyilván itt a legnagyobb az igény az
adatfeldolgozásra és a legtöbb ingerület is
itt keletkezik. Milyen magyarázatot találtak az
értelmes tervezés hívei a sárgafolt
felépítésére? Miért kell az
idegsejteknek szabadon hagyni a fényérzékeny
sejteket éppen azon a ponton, ahol éles képet és
színeket is látunk, ha az emberi szem azért
inverz szem, mert másképpen nem adhatna ilyen minőségű
képet? Mi ennek az ésszerű indoka? A boncolgatott
cikkben miért nem említik meg az emberi szemnek ezt a
meglehetősen fontos jellegzetességét, ha már
a polip szemének hasonló területét
részletesen tárgyalják? Első pillantásra
látszik, hogy ez egy értelmesen tervezett szerv?
Ezen a
területen ugyanis az idegsejtek mintegy "elhúzódnak"
a fényérzékelő sejtek fölül, így
téve lehetővé az éles látást.
Itt azért felmerül a kérdés, hogy az emberi
szem miért inverz szem, ha azon a ponton, ahol az éles
kép keletkezik, az idegsejtek kénytelenek elhajolni a
fényérzékeny sejtek fölül? Az
idegsejtek léte valóban szükséges a szem
működéséhez, ők továbbítják
a fényérzékeny sejtekben keletkező
ingerületet, de akkor miért éppen arról a
területről húzódnak el (természetesen
nem hiányoznak, a látógödör körüli
idegsejtek nyúlványai továbbítják
az itt található fényérzékeny
sejtek ingerületét), ahol a legjobban kellenének,
hiszen nyilván itt a legnagyobb az igény az
adatfeldolgozásra és a legtöbb ingerület is
itt keletkezik. Milyen magyarázatot találtak az
értelmes tervezés hívei a sárgafolt
felépítésére? Miért kell az
idegsejteknek szabadon hagyni a fényérzékeny
sejteket éppen azon a ponton, ahol éles képet és
színeket is látunk, ha az emberi szem azért
inverz szem, mert másképpen nem adhatna ilyen minőségű
képet? Mi ennek az ésszerű indoka? A boncolgatott
cikkben miért nem említik meg az emberi szemnek ezt a
meglehetősen fontos jellegzetességét, ha már
a polip szemének hasonló területét
részletesen tárgyalják? Első pillantásra
látszik, hogy ez egy értelmesen tervezett szerv?
Vizsgáljunk meg egy másik példát is! Léteznek vak emlősök is, például itt a földikutya, vagy Spalax ehrenbergi. Ez egy vakondszerű életmódot folytató, föld alatt élő jószág, amely a közel-keleten elterjedt és a mi szempontunkból azért érdekes, mert bár a genomját nem szekvenálták meg, némi molekuláris munkát azért végeztek rajta. A földikutyák szeme elcsökevényesedett, valahogy úgy, mint a vakondé, ám ez a folyamat jó huszonötmillió éve lejátszódott. Azonban a földikutyák nem élnek teljesen fény nélkül: Bár a szemükkel nem látnak, azért a nap fényéhez igazítják az életüket, így bár képalkotásra nem alkalmas a szemük, azért a fény erősségét érzékeli. Maga ez a szem egy milliméternél valamivel kisebb, láthatóan csökevényes szerv, ami ráadásul egy bőrredő alatt található. A jószág agyában a látókéreg jelentősen csökkent méretű, élettani vizsgálatok semmilyen nyomát sem találták látásnak, ám azt feltárták, hogy a földikutya a biológiai óráját a nappali fényhez és az éjszakai sötétséghez igazítja. Ezt egyszerűen igazolták: Ha éjjel világították meg a földikutyákat és nappal elsötétítették őket, akkor hamar átszoktak erre az életmódra, szokásos nappali tevékenységüket éjjel végezték. Azonban ha a szemüket eltávolították, ez az alkalmazkodóképességük eltűnt. Ezzel összhangban a csökevényes szemből kiinduló idegrostok az agyban található szuprakiazmatikus magba futnak be, amely terület az alvás-ébrenlét körforgását szabályozza emlősökben.
Jannsen J. W. H. és munkatársai 2000–ben a földikutya fényérzékelő szervét vizsgálták. Minden csökevényessége ellenére a földikutya retinája csap- és pálcika sejteket is tartalmaz, ami meglepő bonyolultság, ahhoz képest, hogy nem lát vele. A genomja egy rodopszint kódoló gént is tartalmaz, amely által kódolt fehérje 91-95% -ban hasonló más, látó emlősök rodopszin fehérjéjéhez, sőt, hasonlóan is működik mint más emlősök rodopszinja. Ezek után megvizsgálták a működését a földikutya szemében és azt találták, hogy ott is fényt érzékel. Mit jelent mindez? A nappalok-éjszakák hossza sok emlős életében fontos, mivel ezen adat segítségével követik az évszakokat, a nappalok időtartamához igazítják téli álmukat, szaporodási időszakukat, vagyis életük számos fontos mozzanatát. Viszont a földikutya ehhez egy meglepően bonyolult szervet használ, amely az emlősök szemének elcsökevényesedett változata. Itt éppen annak a példáját látjuk, amikor egy szerv másikká alakul át, ám szerkezetében, működésében még felismerhető eredeti szerepe, ősi formája. Hogyan történhet mindez? Az evolúció alapján adható magyarázat egyszerű: a föld alatt a látás szerepe lecsökkent, így a szem is fölöslegessé vált, ám azon egyetlen működése, amire a köznapi életben nem sokat gondolunk mégis megmaradt, mert az adott állatnak ez fontos. Az új szerkezetű szerv a meglévő, emlősökre jellemző szem változásával alakult ki, ezért hasonlít annyira a földikutya fényérzékelő szerve a többi emlős látó szeméhez. Erre mi az értelmes tervezés talaján nyugvó magyarázat? Miért hasonlít a földikutya fényérzékelő szerve ennyire a többi emlős szemére, ha nyilvánvalóan nem lát vele? Miért nem kapott a feltételezett értelmes tervezőtől egy néhány sejtes szemfoltot, mint amilyen például a laposférgeknek van? És a másik, nem kevésbé fontos kérdés, hogy akkor talán az emlős szem mégsem egyszerűsíthetetlenül összetett? Egyes elemei önmagukban is hasznosak és működőképesek lehetnek...?
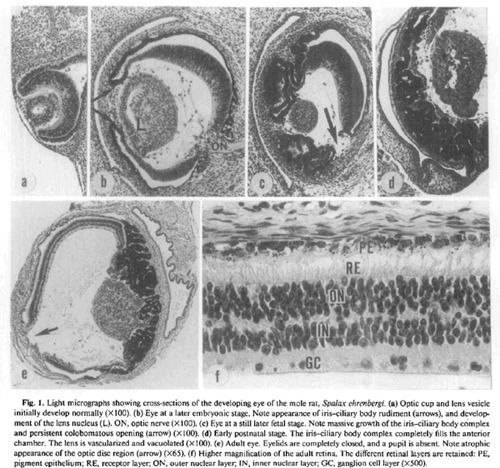 Sanyal S.
és munkatársai 1990. íródott cikkében
szemléletes ábra található a földikutya
szemének fejlődéséről. A szem a látó
emlősökéhez hasonlóan kezd fejlődni, a
korai szem (a) minden szempontból egy tíznapos
egérembrióéhoz hasonló. Megjelenik a
szemlencse (L), és a (b) ábrán láthatóak
a szivárványhártya és a sugártest
kezdeményei is (nyilak). Majd ezek után az embrió
szemének fejlődése teljesen más irányt
vesz: a szemlencse mérete lecsökken, gyakran vérerek
nyomulnak bele, átlátszatlanná válik, a
szem mérete messze nem éri el a hasonló
testtömegű, látó gerincesek szemméretét,
a fejlődésnek indult szemlencse a szemfenékre
tapad, ezzel a más gerincesek szemében betöltött szerepére alkalmatlanná válik. Az (f) kép a földikutya retináját
mutatja, amelyben tisztán felismerhető a látó
emlősök szemére jellemző szerveződés.
PE – az annyit emlegetett pigmentréteg; RE –
fényérzékelő sejtek; ON, IN GC – a
retina idegsejtjei. Ha megfigyeljük, a földikutya szeme is
inverz szem, mint az összes többi emlősé, itt
is tisztán felismerhetőek a fényérzékeny
sejtek és a beérkező fény között
található idegsejtek. Hogy kicsit visszautaljak a cikk
elejére: neki nem teljesen mindegy? Maga az állat vak,
az ő fényérzékeny sejtjei akárhogyan
állhatnának, mégis tisztán felismerhető
az összes többi emlős szemére is jellemző
szerkezet. Ennek a magyarázata az evolúció
talaján maradva egyszerű: egy, a többi emlőséhez
hasonló szem elcsökevényesedésével
alakult ki ez a fényérzékelő szerv, így
bár eredeti feladatát már nem tölti be,
alapszerkezete mégis tisztán felismerhető. Mint korábban már említettem, itt
éppen megfigyelhető egy új szerv kialakulása
egy már meglévőből. Mi az értelmes
tervezés talaján nyugvó magyarázat? Első
pillantásra látszik hogy ez egy értelmesen
tervezett szerv? Ez egy egyszerűsíthetetlenül
összetett szerv? Például miért alakul ki a
szemlencse, ha láthatóan nem működik? Miért
vannak meg a színlátás elemei is ebben a
csökevényes szemben, ha csak a fény erősségét
érzékeli? Miért azonos a földikutya és
az ember retinájának alapszerkezete? (Szerk. megj: részben hasonló szem-sorvadás zajlik a mexikói vaklazacban is.)
Sanyal S.
és munkatársai 1990. íródott cikkében
szemléletes ábra található a földikutya
szemének fejlődéséről. A szem a látó
emlősökéhez hasonlóan kezd fejlődni, a
korai szem (a) minden szempontból egy tíznapos
egérembrióéhoz hasonló. Megjelenik a
szemlencse (L), és a (b) ábrán láthatóak
a szivárványhártya és a sugártest
kezdeményei is (nyilak). Majd ezek után az embrió
szemének fejlődése teljesen más irányt
vesz: a szemlencse mérete lecsökken, gyakran vérerek
nyomulnak bele, átlátszatlanná válik, a
szem mérete messze nem éri el a hasonló
testtömegű, látó gerincesek szemméretét,
a fejlődésnek indult szemlencse a szemfenékre
tapad, ezzel a más gerincesek szemében betöltött szerepére alkalmatlanná válik. Az (f) kép a földikutya retináját
mutatja, amelyben tisztán felismerhető a látó
emlősök szemére jellemző szerveződés.
PE – az annyit emlegetett pigmentréteg; RE –
fényérzékelő sejtek; ON, IN GC – a
retina idegsejtjei. Ha megfigyeljük, a földikutya szeme is
inverz szem, mint az összes többi emlősé, itt
is tisztán felismerhetőek a fényérzékeny
sejtek és a beérkező fény között
található idegsejtek. Hogy kicsit visszautaljak a cikk
elejére: neki nem teljesen mindegy? Maga az állat vak,
az ő fényérzékeny sejtjei akárhogyan
állhatnának, mégis tisztán felismerhető
az összes többi emlős szemére is jellemző
szerkezet. Ennek a magyarázata az evolúció
talaján maradva egyszerű: egy, a többi emlőséhez
hasonló szem elcsökevényesedésével
alakult ki ez a fényérzékelő szerv, így
bár eredeti feladatát már nem tölti be,
alapszerkezete mégis tisztán felismerhető. Mint korábban már említettem, itt
éppen megfigyelhető egy új szerv kialakulása
egy már meglévőből. Mi az értelmes
tervezés talaján nyugvó magyarázat? Első
pillantásra látszik hogy ez egy értelmesen
tervezett szerv? Ez egy egyszerűsíthetetlenül
összetett szerv? Például miért alakul ki a
szemlencse, ha láthatóan nem működik? Miért
vannak meg a színlátás elemei is ebben a
csökevényes szemben, ha csak a fény erősségét
érzékeli? Miért azonos a földikutya és
az ember retinájának alapszerkezete? (Szerk. megj: részben hasonló szem-sorvadás zajlik a mexikói vaklazacban is.)
Kijelenthetjük, hogy értelmes tervezésnek ez esetben sem találtuk nyomát. A puhatestűek egyforma alapszerkezetű szemmel rendelkeznek, nyilván azért, mert az ő közös ősüknek everz szeme volt, ez módosult az egyes leszármazási vonalak mentén a természetes szelekció nyomásának megfelelően. A gerincesek szeme is azonos szerkezetű, még ott is, ahol semmi köze sincs a látáshoz, nyilván az ő esetükben is evolúciós folyamatok működtek, az azonos őstől származó élőlények szeme itt is hasonló. Ellenben a két szem alapszerkezete különbözik egymástól, a két csoport közti átfedéseket nem ismerünk. Egyetlen puhatestűnek sincs olyan szeme mint egy emlősnek és fordítva, egyetlen emlős sem rendelkezik a puhatestűek szeméhez hasonló látószervvel. Kijelenthetjük, hogy az értelmes tervezés elmélete erre az elkülönülésre, vagyis, hogy a feltételezett értelmes tervező miért tartja magát a leszármazási vonalakhoz semmilyen magyarázatot sem ad, ellenben az evolúció talaján maradva mindez érthető és könnyedén magyarázható.
A cikk fejtegetései, miszerint az emberi szem egyes elemeinek sebészi úton való eltávolítása vagy módosítása működésképtelenné tenné mindössze az alapkérdés teljes félreértelmezése. Mint láttuk, a legfontosabb kérdéseket nem is érinti és tulajdonképpen mit bizonyít mindez? Ha egy embernek levágnánk mindkét karját és lábát, könnyen belátható, hogy jelentős hátrányt szenvedne, bizonyára alulmaradna a létért folyó versengésben egy karokkal-lábakkal rendelkező emberrel szemben. De akkor ez azt jelenti, hogy megcáfoltuk a végtag nélküli gerincesek pl. a kígyók és a lábatlan gyíkok létezését...?
Zárszóként elmondható, hogy a kérdés nem az, hogy az emberi szem tökéletes–e, vagy hogy mit kellene rajta változtatni. Az emberi szem olyan, amilyen, amire használjuk, arra tökéletesen megfelel. Tudomásom szerint egyetlen biológus sem fanyalog azon, hogy milyen rossz is az ő szeme, vagy hogy meg akarná műttetni. A kérdés az, hogy az emberi szem felépítéséből következtetni lehet –e az eredetére?
http://en.wikipedia.org/wiki/Compound_eye#Compound_eyes
http://en.wikipedia.org/wiki/Macula
http://www.neurocomputing.org/FrogEye.aspx
Janssen J.
W. H., Bovee-Geurts P. H. M., Peeters Z. P. A., Bowmaker J. K.,
Cooper H.
M., David-Gray Z. K., Nevoi E., DeGrip W. J. (2000) A Fully
Functional Rod Visual Pigment in a Blind Mammal. J Biol Chem 275(49)
Sanyal S.,
Jansen H. G., de Grip W. J., Nevo E., de Jongt W. W. (1990)
The Eye of the Blind Mole Rot, Spalax ehrenbergi
Rudiment With Hidden Function? Invest Ophthalmol Vis Sci 31(7): 1398-1404.

Phasellus lacinia porta ante, a mollis risus et. ac varius odio. Nunc at est massa. Integer nis gravida libero dui, eget cursus erat iaculis ut. Proin a nisi bibendum, bibendum purus id, ultrices nisi.
Utolsó kommentek