
 A fehér ló regék és mítoszok visszatérő szereplője, mesebeli királyfik és valódi kormányzók választott hátasa. Pedig a fehér ló is csak ló, egyetlen különlegessége színében rejlik.
A fehér ló regék és mítoszok visszatérő szereplője, mesebeli királyfik és valódi kormányzók választott hátasa. Pedig a fehér ló is csak ló, egyetlen különlegessége színében rejlik.
Hogy mi okozza ezt a színt, pontosabban színtelenséget, eddig kicsit rejtélyes volt, bár ahogy egyre többet tudtunk meg a pigmentáció biológiájáról, egyre közelebb kerültünk a megoldáshoz (na, meg persze az sem ártott, hogy a genom szekvenálás elterjedésével egyre egyszerűbb és gyorsabb lett viszonylag nagyméretű gének megszekvenálása is). Rendszeres olvasóimban már biztos felébredt a sanda gyanú, hogy a poszt csak egy újabb apropót kínál arra, hogy újabb bőrt húzzak le a testszín-meghatározásban kulcsfontosságú Mc1r receptorról, így gyorsan megnyugtatnék mindenkit, hogy (kivételesen) nem erről lesz szó ;-).
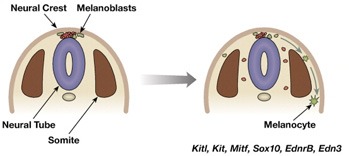 Persze, azért nem is megyünk nagyon messzire. Azokra a sejtekre koncentrálunk, amelyekben aztán az Mc1r kifejti jótékony (sötétpigment-szintetizáló) hatását. Ezek az ún. melanociták, a pigmentsejtek, amelyek fejlődésbiológiailag egy különleges sejtpopulációból származnak. A velőlécet (neural crest) a velőcső (neural tube) összezáródásakor a cső felett levő sejtpopuláció hozza létre. Ezeknek a sejteknek számos fontos feladatuk lesz: pl. a fejben a koponya egyes elemeit, illetve a hozzájuk kapcsolódó izmokat hozzák létre, de részt vesznek a fogak, a pajzsmirigy, a belső fül, a szem és számos ideg kialakulásában is. Ezek mellett szinte mellékesnek tűnik, hogy velőléc eredetű sejtek felelősek a színezet kialakulásáért is.
Persze, azért nem is megyünk nagyon messzire. Azokra a sejtekre koncentrálunk, amelyekben aztán az Mc1r kifejti jótékony (sötétpigment-szintetizáló) hatását. Ezek az ún. melanociták, a pigmentsejtek, amelyek fejlődésbiológiailag egy különleges sejtpopulációból származnak. A velőlécet (neural crest) a velőcső (neural tube) összezáródásakor a cső felett levő sejtpopuláció hozza létre. Ezeknek a sejteknek számos fontos feladatuk lesz: pl. a fejben a koponya egyes elemeit, illetve a hozzájuk kapcsolódó izmokat hozzák létre, de részt vesznek a fogak, a pajzsmirigy, a belső fül, a szem és számos ideg kialakulásában is. Ezek mellett szinte mellékesnek tűnik, hogy velőléc eredetű sejtek felelősek a színezet kialakulásáért is.
A velőcső fölül a lassanként differenciálódó sejtek, így a melanociták is fokozatosan, a testfal mentén lefelé vándorolnak, mígnem el nem érnek végleges helyükre (ezt mutatja a fenti ábra keresztemetszeti ábrázolásban). Ebben a folyamatban (illetve a pigmentsejtek életben maradásában) fontos szerepet játszik egy receptor molekula, amelyet a KIT gén kódol.
Persze előfordulhat az is, hogy a test legtávolabbi pontjaira (ez értelemszerűen a hasi oldalt jelenti van) már nem jutnak el a melanociták, ilyenkor egy jellegzetes fehér "mellény" alakul ki. Ha pedig a melanociták el sem indulnak, vagy menetközben elpusztulnak, akkor egyáltalán nem lesz pigmentáció, testszínezet. S ezzel kvázi le is lőttem a poént, hiszen a jelek szerint ez okozza a fehér lovak fehérségét is. Gyakorlatilag az összes vizsgált esetben (a Franches-Montagnes lovakat, az arab telivéreket, a camarillo fehér és Thoroughbred hátasokat vették górcső alá) a KIT gén különböző mutációira leltek, amelyek vagy működésképtelenné teszik a fehérjét, vagy csökkentik a működőképességét.
Érdekes, bár azért nem meglepő, hogy a lovak nincsenek egyedül ezzel a típusú albinizmussal, hiszen a KIT gént már korábbi vizsgálatok összekapcsolták az emberi piebaldizmus egyes formáival, illetve egér szőrszín mutációkkal is.
Haase B, Brooks SA, Schlumbaum A, Azor PJ, Bailey E, et al. (2007) Allelic Heterogeneity at the Equine KIT Locus in Dominant White (W) Horses. PLoS Genet 3(11): e195 doi:10.1371/journal.pgen.0030195
Steingrímsson E, Copeland NG, Jenkins NA (2006) Mouse coat color mutations: From fancy mice to functional genomics. Dev Dyn 235(9): 2401-2411.
 Hát akkor ismét szőrszín, mert ez (szinte) kifogyhatatlan téma, már csak azért is, mert a laikusok számára is evidens, könnyen érzékelhető változásról van szó.
Hát akkor ismét szőrszín, mert ez (szinte) kifogyhatatlan téma, már csak azért is, mert a laikusok számára is evidens, könnyen érzékelhető változásról van szó.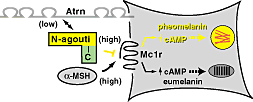 Megnyugtató módon, már a legelején tisztázódott, hogy valami úton-módon, a β-defensin is a klasszikus, Mc1r függő jelátviteli útvonalba szól bele. Sőt, kitartóbb vizsgálódás azt is bizonyította, hogy a fehérje fekete szőrszínt okozó formája erősen tud magához a receptorhoz kötődni, aktivizálva a fekete pigment szintézisét katalizáló folyamatokat. Mindez szép, logikus és kerek, de van egy bökkenő: a β-defensin gén "normál" (nem fekete színt okozó) verziója is képes hasonló erősséggel a receptorhoz kötődni, azaz a kapcsolat puszta ténye önmagában nem lehet a válasz.
Megnyugtató módon, már a legelején tisztázódott, hogy valami úton-módon, a β-defensin is a klasszikus, Mc1r függő jelátviteli útvonalba szól bele. Sőt, kitartóbb vizsgálódás azt is bizonyította, hogy a fehérje fekete szőrszínt okozó formája erősen tud magához a receptorhoz kötődni, aktivizálva a fekete pigment szintézisét katalizáló folyamatokat. Mindez szép, logikus és kerek, de van egy bökkenő: a β-defensin gén "normál" (nem fekete színt okozó) verziója is képes hasonló erősséggel a receptorhoz kötődni, azaz a kapcsolat puszta ténye önmagában nem lehet a válasz. Úgy tűnik, hogy hirtelen nagyon trendi lett neandervölgyi géneket vizsgálni, hiszen
Úgy tűnik, hogy hirtelen nagyon trendi lett neandervölgyi géneket vizsgálni, hiszen 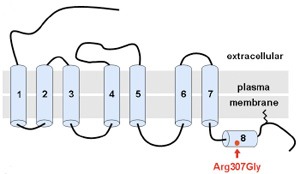 A neandervölgyi mc1r különlegessége, hogy bizonyos esetekben a jelek szerint egy olyan jellegzetes mutációt hordoz(ott), aminek a következtében, működésbe lépésekor a cAMP koncentráció nem éri el azt a szintet, amit a sötét hajú emberekben figyelhetünk meg. Érdekes módon, ez nem azért van (volt), mert a receptor kevésbé hatékony, hanem mert (pontosan nem tisztázott okok miatt) eleve kevesebb mutáns receptorfehérje jut el a sejtek belsejéből a felszínükre. A végeredmény szempontjából azonban ez mindegy és a létrejövő alacsonyabb cAMP szint megegyezik azzal, amit vöröshajú emberekből izolált MC1R hoz létre - azaz jó eséllyel feltételezhető, hogy az ősi neandervölgyiek kb. 1%-a vörös fej- és arcszőrzettel büszkélkedhetett.
A neandervölgyi mc1r különlegessége, hogy bizonyos esetekben a jelek szerint egy olyan jellegzetes mutációt hordoz(ott), aminek a következtében, működésbe lépésekor a cAMP koncentráció nem éri el azt a szintet, amit a sötét hajú emberekben figyelhetünk meg. Érdekes módon, ez nem azért van (volt), mert a receptor kevésbé hatékony, hanem mert (pontosan nem tisztázott okok miatt) eleve kevesebb mutáns receptorfehérje jut el a sejtek belsejéből a felszínükre. A végeredmény szempontjából azonban ez mindegy és a létrejövő alacsonyabb cAMP szint megegyezik azzal, amit vöröshajú emberekből izolált MC1R hoz létre - azaz jó eséllyel feltételezhető, hogy az ősi neandervölgyiek kb. 1%-a vörös fej- és arcszőrzettel büszkélkedhetett. A jelek szerint a történelem valóban ismétli önmagát, különösen ha a szőrszín megváltozásáról van szó.
A jelek szerint a történelem valóban ismétli önmagát, különösen ha a szőrszín megváltozásáról van szó.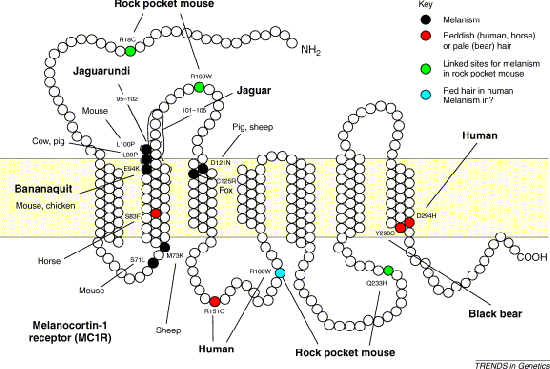


Utolsó kommentek