Ha az emberiség leghosszabb ideje futó és leglátványosabb
eredményeket produkáló genetikai kísérletét kellene megnevezni, akkor a
kutyatenyésztés igen jó eséllyel pályázhatna a lista első helyére,
mint arra az eddigi legteljesebb kutya törzsfát publikáló csapat is
rámutat [1]. Az egyik elterjedt vélekedés szerint az első szelidített
ebek valahol Ázsiában jelentek meg és innen vándoroltak különböző nomád
csoportok oldalán a sarkkör felé, Afrikába illetve végül Európába.
Mindenesetre ezt látszik alátámasztani a fent említett tanulmány
eredménye is, hiszen a kutyák törzsfáján a farkashoz legközelebbi ebek
ősi ázsiai és arktikus fajtákhoz tartoznak (1. Ábra).
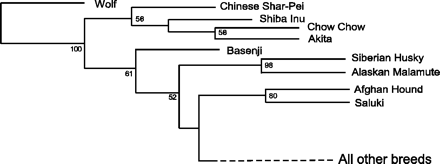 1. Ábra: Nyocvanöt kutyafajta és a szürke farkas genetikai
összehasonlításából készült törzsfa. Az ázsiai és arktikus fajokat
tartalmazzák a legrégebbi elágazások, míg a legtöbb Európában
nemesített faj a klasszikus statisztikai módszerekkel nehezen
szétválasztható "All other breeds" csoportba került. (Ezen belül
érzékenyebb módszerek három nagyobb csoportot különítettek el: az
elsőbe elsősorban a pásztorkutyák pl. skótjuhász, komondor, kuvasz
kerülnek, de ide tartozik a bernáthegyi és a szürke agár is (ez utóbbi
meglepő lehet, hiszen az afgán agár az egyik ősi fajtaként jelenik meg:
arról lehet szó, hogy a szürke agarat később nemesítették az afgán
névrokonától függetlenül), a másodikba a masztif-félék (masztif,
bulldog, boxer, bull-terrier) találhatók (itt külséejében "kakukktojás"
a németjuhász) még minden más fajta a harmadik csoportban van.) [1]
1. Ábra: Nyocvanöt kutyafajta és a szürke farkas genetikai
összehasonlításából készült törzsfa. Az ázsiai és arktikus fajokat
tartalmazzák a legrégebbi elágazások, míg a legtöbb Európában
nemesített faj a klasszikus statisztikai módszerekkel nehezen
szétválasztható "All other breeds" csoportba került. (Ezen belül
érzékenyebb módszerek három nagyobb csoportot különítettek el: az
elsőbe elsősorban a pásztorkutyák pl. skótjuhász, komondor, kuvasz
kerülnek, de ide tartozik a bernáthegyi és a szürke agár is (ez utóbbi
meglepő lehet, hiszen az afgán agár az egyik ősi fajtaként jelenik meg:
arról lehet szó, hogy a szürke agarat később nemesítették az afgán
névrokonától függetlenül), a másodikba a masztif-félék (masztif,
bulldog, boxer, bull-terrier) találhatók (itt külséejében "kakukktojás"
a németjuhász) még minden más fajta a harmadik csoportban van.) [1]
Mindenestre a szorgalmas és céltudatos tenyésztgetésnek meglett az
eredménye mind genetikailag (a fajták közötti genetikai különbség a
különböző kutyák közti variáció közel harmadát teszi ki -
összehasonlításként a nagyon elkülönült humán populációk között is max.
5-10% ez az arány), mind mint azt nyilván fölösleges ecsetelnem
kinézetileg (fenotípusosan). Mi okozhatott ilyen - viszonylag - gyors
fenotípusos változást? Az egyik lehetséges okról, a gének szabályozó
szekvenciáiban bekövetkező változásról már írtam korábban, de van egy másik igen érdekes elképzelhető ok is.
A különböző genomokban viszonylag gyakoriak az ún. mikroszatelliták.
Ezek általában két vagy három bázis ismétlődéséből állnak (pl.
CACACACACACA... vagy CAGCAGCAGCAGCAG...), előbbi főként nem kódoló
DNS-re jellemző, míg utóbbi a fehérjéket kódoló DNS-re . Mindmáig nem
teljesen tisztázott okokból, de a legvallószínűbbnek a DNS polimeráz
"megcsúszását" tartják, az ismétlődések száma gyakran változik
különböző egyedek között (ez az oka annak, hogy kódoló szekvenciában
triplet ismétlődések vannak, hiszen a genetikai kód is "hárombetűs",
így a csúszás erdeménye egy extra aminosav, míg dinukleotida ismétlődés
esetén, az egész "leolvasó keret" eltolódna és a mikroszatellita után
levő DNS teljesen mást kódolna). Ezt egyrészt remekül lehet
hasznosítani rokonsági vizsgálatokban (ha közeli rokonokban vizsgálunk
több mikroszatellitát, azok nagy valószínűséggel egyezni fognak),
másrészt azonban a túl hosszúvá váló ismétlődések olyan súlyos
betegségeket okozhatnak mint a spinocerebelláris ataxia vagy a Huntington-kór.
Egy texasi kutatócsoport azt kezdte el vizsgálni, hogy összefüggésbe
hozhatók-e a mikroszatelliták hossza az egyes kutyák morfológiai
tulajdonságaival [2].
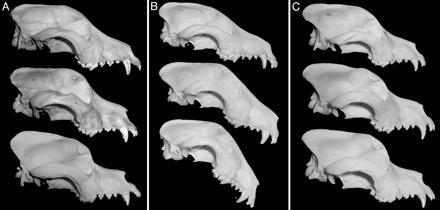 2. Ábra: Kutyakoponyák morfológiájának gyors
változása: az A panelben fajtiszta bernáthegyi koponyák láthatók
~1850-ből (felül), 1921-ből (középen) és 1967-ből (alul), a B panelen
bull-terrier koponyák 1931-ből, 1950-ből és 1967-ből, végül a C panelen
újfullandi koponyák 1926-ból, 1964-ből és 1971-ből. A középső
terrier-koponya gazdájának runx-2 génjében az egyik
mikroszatellita hosszabb mint a modern bull-terrierek azonos génjében.
(Ez alátámasztja a kutatók modelljét, de azért egy egyfős minta nem
igazi, szóval csak óvatosan lelkesedni... ;-)) [2]
2. Ábra: Kutyakoponyák morfológiájának gyors
változása: az A panelben fajtiszta bernáthegyi koponyák láthatók
~1850-ből (felül), 1921-ből (középen) és 1967-ből (alul), a B panelen
bull-terrier koponyák 1931-ből, 1950-ből és 1967-ből, végül a C panelen
újfullandi koponyák 1926-ból, 1964-ből és 1971-ből. A középső
terrier-koponya gazdájának runx-2 génjében az egyik
mikroszatellita hosszabb mint a modern bull-terrierek azonos génjében.
(Ez alátámasztja a kutatók modelljét, de azért egy egyfős minta nem
igazi, szóval csak óvatosan lelkesedni... ;-)) [2]
A koponyamorfológiát befolyásoló transzkripciós faktorokat
végignézve korellációra leltek a Runx-2 nevű fehérjében levő
mikroszatelliták hossza és a koponya alakja között. A Runx-2-ben
kétfajta mikroszatellita van egymás után: az egyik egy poli-glutamin
(polyQ) szekvenciát kódol, a másik pedig egy poli-alanint (polyA).
Általában más kísérleti rendszerekben a polyQ növekedés (bizonyos
határok között) a transzkripció aktivációs képességet növelik, míg a
polyA növekedés ugyanazt csökkenti. A kimutatott összefüggés szerint a
polyQ/polyA arány összefügg a koponya hosszával és hajlásával (2. Ábra).
Hogy egy kis plusz zamata legyen a sztorinak a pireneusi
juhászkutyákban jelen levő polidaktíliát (hat lábujjuk van az ebeknek)
is hasonló okra tudták visszavezetni. Ezúttal az Alx4 transzkripciós
faktorban leltek egy 51 bázispárnyi deléciót (amely semelyik más
kutyafajtában nem volt jelen). Ennek okán valószínűleg az Alx4
működésképtelenné válik és nem tudja a célgénjeit aktiválni a fejlődő
végtagbimbóban; az Alx4 mutáns allélját hordozó egerek is egy extra
lábujj büszke tulajodnosai (3. Ábra).
3. Ábra: Az Alx-4 mutációja mind egerekben, min pireneusi juhászkutyákban többujjúságot (polidaktíliát) okoz. [2]
[1] Parker HG, Kim LV, Sutter NB, Carlson S, Lorentzen TD, Malek TB, Johnson GS, DeFrance HB, Ostrander EA, Kruglyak L. (2004) Genetic structure of the purebred domestic dog. Science 304:1160-1164.
[2] Fondon JW 3rd, Garner HR. (2004) Molecular origins of rapid and continuous morphological evolution. Proc Natl Acad Sci USA 101(52):18058-63. Epub 2004 Dec 13.


Utolsó kommentek