
 Pontosítás végett nem fosszíliákban levő génekről lesz szó (bár az is nagyon érdekes), hanem olyan DNS szakaszokról, amelyek nagyon is hús-vér élőlényekben fordulnak elő. S bár szekvenciájuk, ill. szerkezetük alapján könnyedén azonosítható, hogy ezek milyen gének lennének, a bennük levő, évmilliók alatt felgyülemlett mutációk működésképtelenné tették őket, így (ha naggyon költői akarok lenni) ma már csak önmaguk árnyékaként tengődnek a kromoszómákon. Ezeket a nukleinsav darabokat nevezzük hát "fosszilis"-, vagy egyszerűbben pszeudogéneknek.
Pontosítás végett nem fosszíliákban levő génekről lesz szó (bár az is nagyon érdekes), hanem olyan DNS szakaszokról, amelyek nagyon is hús-vér élőlényekben fordulnak elő. S bár szekvenciájuk, ill. szerkezetük alapján könnyedén azonosítható, hogy ezek milyen gének lennének, a bennük levő, évmilliók alatt felgyülemlett mutációk működésképtelenné tették őket, így (ha naggyon költői akarok lenni) ma már csak önmaguk árnyékaként tengődnek a kromoszómákon. Ezeket a nukleinsav darabokat nevezzük hát "fosszilis"-, vagy egyszerűbben pszeudogéneknek.
Mivel a pszeudogének (talán fölösleges is hangsúlyozni) értelemszerűen egy távoli ősben nagyon is működőképesek voltak, puszta létük egyszerre két fontos dolgot is bizonyít: az élőlények közös leszármazását (hiszen egy "intelligensen tervezett" szervezetbe, minek tennénk működésképtelen génszakaszokat...?), valamint azt, hogy csak azok a DNS szakaszaink mentesek a változástól, amelyek fontosak és megváltozásuk az élőlényt igencsak hátrányosan érintené. Egyébként, "ami elromolhat, az el is romlik".
A fent megfogalmazottak persze azt is jelentik, hogy pszeudogének akkor jöhetnek létre, amikor egy-egy specifikus sejtcsoport/szövet/szerv elveszti a funkcióját. A legjobb példa erre, gondolhatnánk józan paraszti ésszel, a csökevény szervek lennének, de azért a helyzet ennél bonyolultabb. Például kígyók esetében a végtagelvesztést nem kísérte a végtagfejlődésben szereplő gének pszeudogénesedése, hiszen ezek a gének (illetve termékeik) még számtalan más funkcióval rendelkeznek, így nem nélkülözhetőek. Max. a végtagspecifikus szabályozószekvenciáik degenerálódhattak, ami persze a mi sztorink szempontjából közel sem ugyanaz.
Azaz pszeudogének vadászata közben olyan génekre kell leszűkítsük a keresőképünket, amelyek igen specifikusan csak egy-egy funkcióval rendelkeznek, és ez a funkció egyes fajokban megszűnt. Ez nagyon rejtélyesen hangozhat, így inkább gyorsan ugorjunk konkrét példákra.
1.) A sziláscetek legnevezetesebb "része" a szájukban lógó és a plankton kiszűrését végző szila-együttes. Persze nemcsak az a jellegzetes, ami van nekik, hanem az is, ami nincs: ez pedig (a hátsó végtag mellett) egy rendes fogsor. Bár a szilák kialakulása evolúciósan ill. fejlődéstani szempontból még nem teljesen tisztázott (ami nem csoda, hiszen finoman szólva sem triviális szürkebálna embriókat beszerezni...), , a szilásceteket összevetve közeli rokonaikkal, az ámbrás cetekkel, vagy delfinekkel, kézenfekvő azt a következtetést levonni, hogy ez a folyamat "mellesleg" a fogazat elvesztésével járt. Ez persze nem váratlan, hiszen foggal nem lehet kiszűrni a planktont, így aztán elvesztése sem jelenthetett túl nagy veszteséget a sziláscet-ősnek (feltéve, persze, ha már volt szilája...).
Mindenesetre a fogfejlődésben van néhány nagyon specifikus gén, ami csak ebben a folyamatban szerepel, pl a zománc kialakításában is részt vevő enamelin és ameloblasztin. Mint arra nemrég fény derült, mindkét fehérjét kódoló gén (ENAM és AMBN) pszeudogénesedett a szilascetek csoportjában: szekvenciájuk az fellelhető, de a mutációk miatt arról működőképes fehérje már nem jöhet létre. Ezzel szemben, a fogfejlődésen kívül még más funkciókat is betöltő DMT1 gén (dental matrix protein 1) "él" és virul, pont ahogy a felvezetőben vázolt logika diktálná.
2.) A jéghideg antarktiszi vizekben élő különleges jéghalak talán legfurcsább tulajdonsága, hogy fehér a vérük. Mindez annak "köszönhető", hogy nincsenek vörösvérsejtjeik, és szöveteik a környező oxigéndús vizekből beoldódó oxigént használják anyagcsere folyamataikhoz. Így persze a más fajokban a gáz molekulák szállítására szakosodott hemoglobint alkotó, α- és β-globin molekulák is elvesztették a fontosságukat - és mint az gondolom már nem annyira meglepő, az őket kódoló gének is "fosszilizálódtak".
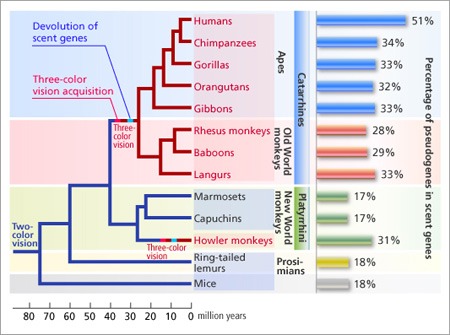 3.) Az ember és emberszabású majmok trikromatikus látása az élővilág teljes egészében nem, de az emlősök közt annál inkább különlegesnek számít. Mégpedig azért, mert az evolúciójuk hajnalán az emlősök (feltehetőleg az éjszakai életmódhoz való alkalmazkodás során) elvesztették több opszin génjüket is, s így aztán egy kutya vagy egy egér csak dikromatikus látásra képes (vagyis az általa érzékelt színskála csak vörösből és kékből "keveredik" ki).
3.) Az ember és emberszabású majmok trikromatikus látása az élővilág teljes egészében nem, de az emlősök közt annál inkább különlegesnek számít. Mégpedig azért, mert az evolúciójuk hajnalán az emlősök (feltehetőleg az éjszakai életmódhoz való alkalmazkodás során) elvesztették több opszin génjüket is, s így aztán egy kutya vagy egy egér csak dikromatikus látásra képes (vagyis az általa érzékelt színskála csak vörösből és kékből "keveredik" ki).
Az emberszabású majmok kialakulása közben az egyik megmaradt opszin-gén duplikációja , majd változása következtében, kialakul egy zöld fényt is érzékelő opszin, így aztán mi a színskála lényegesen szélesebb spektrumával barátkozhatunk életünk során.
Ugyanakkor nem biztos, hogy az ebből a szempontból kevésbé "szerencsés" emlősök sokkal kevesebb információval rendelkeznek a világról: hiszen velünk ellentétben lényegesen jobb a szaglásuk. Illetve talán az a helyesebb, ha azt mondom, hogy a mienk lényegesen rosszabb: mert míg ugyanúgy kb. 1000 szag-receptor gént hordozunk a genomunkban, mint egy eb, a mi esetünkben ezek több mint fele pszeudogén, míg négylábú barátaink esetében ez az arány kevesebb mint 20%.
A két jelenség (trikromatikus látás kialakulása ill. szagreceptor-gének elsatnyulása) minden valószínűség szerint összefügg. A jobb látás kicsit fölöslegessé tette a hatalmas szagreceptor repetoárt, és így abból egy-két gén elvesztése nem érintette nagyon súlyosan az emberszabású őst, aki amúgy szín alapján is ki tudta már szúrni az érett gyümölcsöt.
Persze lehetünk szkeptikusak (sosem árt ;-)), és felvethetjük, hogy a két jelenség együttes megjelenése még nem kell ok-okozati viszonyt feltételezzen. Ennek a megvizsgálására az újvilági majmokhoz fordulhatunk: ezek többsége dikromatikus látással rendelkezik, egyetlen, annál árulkodóbb kivétellel, ez pedig a bőgőmajom (howler monkey). A bőgőmajmoknak ugyanis szintén trikromatikus látáshoz van szerencséjük, és amikor "belenéztek" a szagreceptoraikba, az derült ki, hogy azok lényegesen nagyobb százaléka működésképtelen, mint a közeli rokonaikban. Vagyis úgy tűnik, hogy többről van szó egyszerű egybeesésnél: az éles látás kiváltja az éles szaglást - mármint egy egy főemlős hétköznapjaiban.
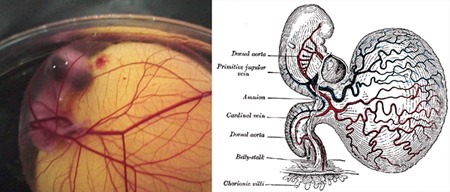 4.) Az emlős (és így természetesen emberi) embriók érdekes tulajdonsága, hogy korai fázisukban egy-egy jellegzetes szikzacskóhoz kapcsolódnak (lásd mellékelt ábra jobboldali panelje). Ez a szerv igen fontos, hiszen a falaiban futó erekben zajlik az embrionális vérképződés első hulláma, ugyanakkor nem lehet nem észrevenni, hogy ez mégiscsak egy üres zsák.
4.) Az emlős (és így természetesen emberi) embriók érdekes tulajdonsága, hogy korai fázisukban egy-egy jellegzetes szikzacskóhoz kapcsolódnak (lásd mellékelt ábra jobboldali panelje). Ez a szerv igen fontos, hiszen a falaiban futó erekben zajlik az embrionális vérképződés első hulláma, ugyanakkor nem lehet nem észrevenni, hogy ez mégiscsak egy üres zsák.
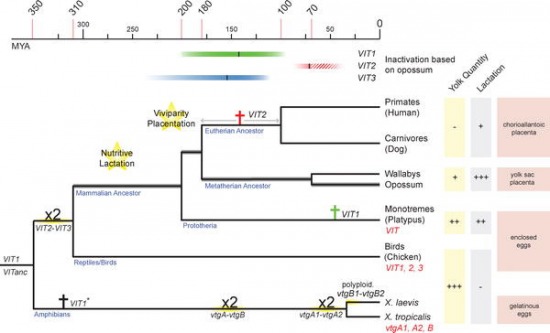
Evolúciósan nézve ez messze nem volt mindig így, elég egy hüllő, vagy egy csirke embrióra (bal oldali panel) vetni egy pillantást: a tojással szaporodó fajokban a szikzacskóban a szikanyag található (na vajon honnan jött a neve ... ;-)), magyarán a tojás sárgája. Ez az a tápanyag, amiből az embrió fedezi kezdeti energiaszükségletét és lezavarja azt a rengeteg sejtosztódást, amelyek végén egy kis csirke bújhat ki a tojásból. Az emlősök, különösen a méhlepényesek és kisebb mértékben az erszényesek is, azonban új trükkökkel álltak elő. Ezek egyike a placenta, vagyis a méhlepény, amelyen keresztül az embrió kezdeti fejlődése során közvetlenül a szülő szervezetéből juthat a táplálékhoz, a másik pedig az anyatej, ami születés után lesz hosszabb ideig az elsődleges élelemforrás. És bár ezek elsődlegesen nem a szikanyag kiváltására "jöttek létre", hosszabb távon mégis ehhez vezettek.
A szikanyag fölöslegessé válása azzal is járt, hogy a szikanyagot kódoló ún. vitellogenin gének (VIT) - amelyekből az emlősök és madarak ill. hüllők közös ősében három is volt - lassanként pszeudogénekké váltak. Érdekes módon a folyamat viszonylag lassan ment végbe, olyannyira hogy minimum egy-két gén inaktiválódására a különböző emlős vonalakban (erszényes vs. méhlepényes), azok szétválása után párhuzamosan került sor.
Nade, hopp, szemérmesen hallgattam eddig a legkülönlegesebb emlősökről, a tojással szaporodó kloákásokról, ahova a mindenki kedvence kacsacsőrű emlős is tartozik. Ha a méhlepény léte váltotta ki a szikanyagot (ill. a VIT gének működőképességét), akkor mi is a helyzet velük?
Nos ez esetben, talán nem meglepő módon, az egyik VIT gén még minden jel szerint működőképes. Így aztán, bár a gén két társa mára már csak a "díszként" ül a genomban, annyi szikanyagot még képes az anya állat termelni, hogy a utódjait fejlődésük kezdeti szakaszán túllendítse. Aztán pedig itt is jöhet a már említett anyatej.
(Az illusztrációként felhasznált képek innen származnak: Nikon, Freshpics, Wikipedia.)
Brawand D, Wahli W, Kaessmann H (2008) Loss of Egg Yolk Genes in Mammals and the Origin of Lactation and Placentation. PLoS Biol 6(3): e63 doi:10.1371/journal.pbio.0060063
Deméré TA, McGowen MR, Berta A, Gatesy J (2008) Morphological and molecular evidence for a stepwise evolutionary transition from teeth to baleen in mysticete whales. Syst Biol 57(1): 15-37.
Near TJ, Parker SK, Detrich HW 3rd. (2006) A genomic fossil reveals key steps in hemoglobin loss by the antarctic icefishes. Mol Biol Evol 23(11): 2008-2016. doi:10.1093/molbev/msl071
Glusman G, Yanai I, Rubin I, Lancet D (2001) The complete human olfactory subgenome. Genome Res 11(5): 685-702.
Gilad Y, Wiebe V, Przeworski M, Lancet D, Pääbo S (2004) Loss of Olfactory Receptor Genes Coincides with the Acquisition of Full Trichromatic Vision in Primates. PLoS Biol 2(1): e5. doi:10.1371/journal.pbio.0020005

{kind=link}
Utolsó kommentek