
 A kínai lelőhelyek továbbra is ontják magukból az érdekesnél érdekesebb fosszíliákat. Most éppen ismét egy ősi emlős került elő, amely egyszerre támasztja alá a középfül evolúciós kialakulásáról szóló elméleteinket és kínál betekintést az evolúció boszorkánykonyhájának egy Hox-gének tüzelte bugyrába.
A kínai lelőhelyek továbbra is ontják magukból az érdekesnél érdekesebb fosszíliákat. Most éppen ismét egy ősi emlős került elő, amely egyszerre támasztja alá a középfül evolúciós kialakulásáról szóló elméleteinket és kínál betekintést az evolúció boszorkánykonyhájának egy Hox-gének tüzelte bugyrába.
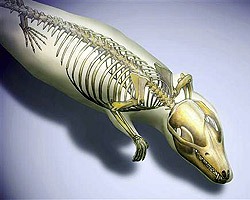
A Yanoconodon allini (mert ez a neve nekije) jellegzetes fogazata alapján a virágkorát a mezozoikumban elérő, menyétszerű állatokat magába foglaló Eutriconodonta csoportba tartozik.
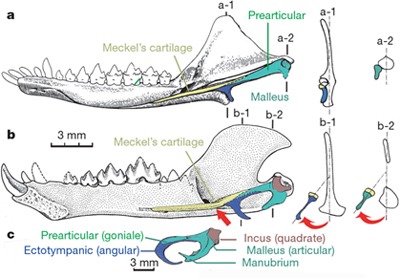
Ami azonban igazán érdekes a leletben, az a füle, pontosabban azoknak a csontocskáknak a helye és alakja, amelyek a mi középfülünkben a kalapács (maleus) - üllő (incus) - kengyel (stapes) trimuvirátust alkotják. Ezek közül az első kettő az első embrionális kopoltyúívből keletkezik (részben az embrionális korban jelen levő Meckel-porc csontosodásával), míg az utóbbi a második kopoltyúív terméke.
A hüllőkben azonban egy kicsit más a helyzet: esetükben az első kopoltyúívből létrejövő csontok kizárólag az állkapocs felépítésében játszanak szerepet, vagyis a kalapács és üllő homológjai is más funkciót töltenek be, mint bennünk emlősökben. Az egyedüli középfül-csont a kengyel, amely a szárazföldhöz való alkalmazkodás során alakult ki.
 A nagy kérdés az tehát, hogy vajon mennyiben követhetjük az ominózous két csontocska átalakulását az emlősök evolúciója során. A válasz már eddig is az volt, hogy elég sokban és a Yanoconodon remekül beillik a sorba.
A nagy kérdés az tehát, hogy vajon mennyiben követhetjük az ominózous két csontocska átalakulását az emlősök evolúciója során. A válasz már eddig is az volt, hogy elég sokban és a Yanoconodon remekül beillik a sorba.
A baloldali ábrán legfelül (a) egy ősi emlős, a Morganucodon alsó állkapcsa látható, alatta pedig (b) a Yanoconodon-é, legalul pedig egy kacsacsőrű emlős fülcsontocskái. Talán nem is kell hosszan ecsetelni, hogy a utóbbi alig különbözik a Yanoconodon homológ csontjaitól, talán csak az utóbbiban teljes egészében csontosodott Meckel porc miatt látszik különbözőnek. Így az embrionális fejlődés során (amikor a Meckel-porc még jelen van) a két koponya még hasonlóbbnak tűnik.
A másik érdekességnek, mint azt előre beharangoztam, a Hox-génekkel való evolúciós kavarás bizonyítéka tűnik. No persze, mivel DNS-t nem tudunk izolálni ezekből az ősi csontokból, a bizonyíték kissé indirekt, de azért nem lebecsülendő.
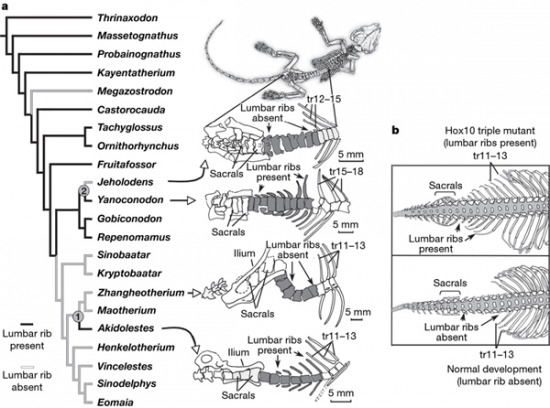
A Hox gének egyik fontos szerepe ui. a csigolyák identitásának kialakítása. A modern emlősök gerincoszlopan hét nyak-, 13-14 hát- és 5-6 ágyékcsigolya lelhető fel. Utóbbi két csoport között az egyik jellegzetes különbség a bordák megléte ill. hiánya, amely a Hox10 gének működésének függvénye (a Hox10 paralógok hiánya egerekben borda fejlődést serkent az ágyékcsigolyákon). A Yanoconodon 15 hát- és 7 ágyékcsigolyával rendelkezett, s utóbbiakon egyértelműen bordák figyelhetőek meg. Ezzel szemben számos olyan korabeli emlősfaj ismert (egyesek a Yanoconodon közeli rokonai, pl. Jeholodens), amelyek híján vannak ezeknek az ékességeknek, vagyis ezek az állatok pont egy olyan korszakból származnak, amikor a természet elkezdett kacérkodni a Hox10-függő borda-represszió gondolatával. S mivel történetesen később egyes ágyékbordát nélkülöző állatok sikeresebbek lettek, mint más társaik (nem feltétlenül a bordák hiánya miatt),ma az ő utódaik népesítik be az élet porondját.
Luo, ZX, Chen, P, Li, G, Chen, M (2007) A new eutriconodont mammal and evolutionary development in early mammals. Nature 446: 288-293.

{kind=link}
Utolsó kommentek