
 Bár az emberi fajt „az evolúció csúcsának“ hirdető nézetek egy erőltetetten és erősen szubjektív emberi álláspontot tükröznek (ti. pusztán evolúciós értelemben minden ma élő faj egyformán sikeresnek tekinthető), az azonban kétségtelen, hogy fajunk különleges. Különleges kommunikációjának és eszközhasználatának bonyolultságában, elvont gondolkozásának mélységében és nem utolsó sorban azért is, mert minden jel szerint bennünk ébredt először az evolúciós folyamat öntudatra bolygónk története során. Mindez pedig elég indokolttá teszi azt a velünk közel egyidős kíváncsiságot, hogy megértsük mi az ami "emberivé" tesz bennünket, létezik-e valami kézzel is megfogható oldala különlegességünknek.
Bár az emberi fajt „az evolúció csúcsának“ hirdető nézetek egy erőltetetten és erősen szubjektív emberi álláspontot tükröznek (ti. pusztán evolúciós értelemben minden ma élő faj egyformán sikeresnek tekinthető), az azonban kétségtelen, hogy fajunk különleges. Különleges kommunikációjának és eszközhasználatának bonyolultságában, elvont gondolkozásának mélységében és nem utolsó sorban azért is, mert minden jel szerint bennünk ébredt először az evolúciós folyamat öntudatra bolygónk története során. Mindez pedig elég indokolttá teszi azt a velünk közel egyidős kíváncsiságot, hogy megértsük mi az ami "emberivé" tesz bennünket, létezik-e valami kézzel is megfogható oldala különlegességünknek.
Filozófiai értekezések hosszú sorát lehetne írni a témáról, de itt és most a probléma egy profánul materialista oldalát próbáljuk megfogni: hogyan kódolódnak génjeinkben emberi vonásaink. Hogy a poént, pontosabban annak nem létét, hamar lelőjem: erre most még (?) nem tudunk felelni, de az elmúlt évek mindenesetre számos eszközt adtak a kezünkbe, amelyek segítségével nagyvonalakban felvázolhatjuk, mi is történhetett az utóbbi 5-7 millió év során, miután a csimpánz- és saját vonalunk szétvált.
A kutatások eddig elsősorban (de nem kizárólag) az idegrendszer fejlődésére koncentráltak, nem véletlenül. A csimpánz - ember összehasonlításban szembetűnő egyes kognitív képességek megugrása, ráadásul az említett pár millió év alatt az emberi-vonalban legradikálisabban éppen az agy mérete változott. S mivel éppen ez a szervünk az összes közül messze a legenergiaigényesebb, ill. megnagyobbodása jelentősen megnehezítette a szülést ill. elhúzta az egész emrbionális és poszt-embrionális fejlődést, a változás egyértelműen hasznos kellett legyen, különben csúnyán fennakadt volna a természetes szelekció érzéketlen rostáján.
Az elmúlt pár évben három igen jelentős adathalmaz került birtokunkba, amelyek nemcsak az emberi idegrendszer, de úgy általában teljes szervezetünk evolúciójának megértését segíthetik. Ezek: az emberi genom, a velünk genetikailag leginkább rokon csimpánzok genomja, illetve (múlt hét óta) egy távolabbi rokon, a makákó genomja. (Utóbbi azért lényeges, mert bár a csimpánz és emberi genomok összehasonlítása önmagában is informatív, pusztán kettejük alapján nehéz választ adni arra, hogy az egyes különbségek melyik vonalon jelentek meg. A makákó genommal összevetve viszont azt is láthatjuk, hogy melyik faj hordozza egyes gének "ősi" allélját, és melyik hordoz egy újkeletűbb variánst.)
A genomprojectek (részleges) befejezése előtt, az a fajta masszív genom-összehasonlítás, ami ma már hétköznapinak számít (és amiről később itt is esik szó), értelemszerűen lehetetlen volt, éppen ezért a kutatások, részben kényszerből, olyan génekre koncentráltak, amelyek mutációi emberekben komoly kognitív változásokat okoztak - vagy az agy méretének befolyásolásával, vagy anélkül. Ha ezen gének emberi és csimpánz homológjait összehasonlítva változásokat találunk, logikusnak látszik felvetni: konkrétan ezen változásoknak köszönhető egy-egy kognitív tulajdonság terén a különlegességünk. A három leghíresebb és legtöbbet tanulmányozott ilyen gén példáján keresztül azonban rögtön látni fogjuk, hogy már a kérdésfelvetés sem annyira logikus, mint amilyennek első pillanatban látszik, az pedig, hogy a fent említett "gyanút" néhány esetben kvázi tényként állították be, mára kifejezetten szar intuíciónak bizonyult.
Az első példánk a FOXP2 lesz, amelyet felfedezése óta szerényen csak "beszédgén"-ként aposztrofál a média. Magát a gént egy olyan angol családban fedezték fel, amelynek sok tagja komoly beszédproblémákkal küszködött. Nemcsak nehezebben artikuláltak, de a nyelvtani szabályokkal is meggyűlt a bajuk, és ennek megfelelően az agyi aktivitást vizsgáló eljárások a beszédben kiemelten fontos Broca-területen csökkent működést mutattak ki. A közelebbi vizsgálatok aztán azt is kimutatták, hogy a jelenség egy transzkripciós faktorban bekövetkezett mutációval hozható kapcsolatba, ez a FOXP2.
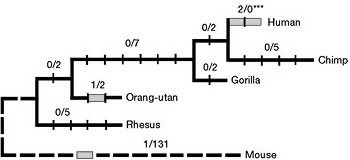 Már az eredeti felfedezést is nagy érdeklődés övezte és erre még sikeresen rátett egy lapáttal a Nature egyik tanulmánya, ami azt mutatta ki, hogy bár az egér és ember FOXP2 aminosav-szekvenciája között összesen három különbség fedezhető fel (szürke téglalapok), ezek közül kettő az ember felé vezető vonalban a csimpánzzal való szétválás után jelent meg. Sőt, a humán verzió az elmúlt százezer év során terjedt el nagy ütemben, ami arra utal, hogy a két változás igencsak adaptív.
Már az eredeti felfedezést is nagy érdeklődés övezte és erre még sikeresen rátett egy lapáttal a Nature egyik tanulmánya, ami azt mutatta ki, hogy bár az egér és ember FOXP2 aminosav-szekvenciája között összesen három különbség fedezhető fel (szürke téglalapok), ezek közül kettő az ember felé vezető vonalban a csimpánzzal való szétválás után jelent meg. Sőt, a humán verzió az elmúlt százezer év során terjedt el nagy ütemben, ami arra utal, hogy a két változás igencsak adaptív.
Bár ezen a ponton logikusnak tűnne a FOXP2 fehérjét ért két evolúciós változást sztárolni, nem árt néhány dolgot a fejünkben tartani. Egyrészt semmi bizonyíték nincs arra, hogy a csimpánz FOXP2 ugyanúgy működésképtelen lenne, mint a hibás emberi fehérje (utóbbiban a mutáció máshova esik, mint az ember-csimpánz különbségek), ez pusztán egy jól hangzó elmélet. Másrészt pedig a FOXP2 nemcsak az agyban fejeződik ki, hanem a tüdőben, szívben és gyomorban is - azt pedig szintén nem bizonyította senki, hogy a FOXP2 szelekciója agyi funkciója miatt következett be. Harmadrészt, az említett két aminosav-változáson kívül még számos nukleotid-szintű mutáció fedezhető fel az emberi FOXP2 génben és az szintén nem zárható ki, hogy ezek egyike egy fontos szabályozó-mutáció, és a szelekció ezt próbálta megőrizni és nem a fehérjekülönbségeket.
Ezeket a fenntartásokat azért emeltem ki, mert néhány fontos kísérlet súlyt ad másfajta megfontolásoknak is. Például a FOXP2 hiányában az egerek kommunikációja/vokalizációja sem működik megfelelően, ami arra utal, hogy a génnek inkább van szerepe a kommunikációban általában, mint konkrétan az emberi beszéd kialakulásában. Emellett énekes- és nem énekesmadarak FOXP2 génje teljesen egyforma, de előbbiekben épp az énektanulás során a gén expressziója feltűnően megnő a folyamatban fontos agyi területen. (Ez ismét amellett szól, hogy nem konkrétan a gén szekvenciája, hanem kifejeződésének helye és ideje az, ami számít.)
A humán betegségek alapján evolúciósan fontos géneket kereső eljárás másik veszélyére a MCPH1 - ASPM génpáros ad jó példát. Ezek mindegyike igen fontos az agyméret kialakulásában (feltehetőleg az idegsejtek osztódását szabályozzák), és gyakori okozói a microcephaliának, vagy kisfejűségnek. Mivel mindkét gén szekvenciája jelentős változásokat élt meg a főemlősök (az ASPM esetében különösen az ember) evolúciója során, szintén remek jelöltek arra, hogy a kognitív képességeink forrását bennük keressük. Ezért is volt bő két éve nagy szenzáció, amikor olyan adatok láttak világot, amelyek arra utaltak, hogy mindkét gén erős szelekció alatt van az emberi populációkban. Találgatások indultak meg, hogy a gének preferált alléljai biztos nagyobb intelligenciát, kreativitást, stb. biztosítanak - de végül egyik sem nyert kísérletes megerősítést. Ennek egyik oka az lehet, hogy akárcsak a FOXP2, ez a két gén sem kizárólag az agyban fejeződik ki, vagyis könnyen lehet, hogy más szervekben betöltött funkcióik okán terjedtek el a preferált variánsok. Különösen igaz ez az ASPM-re, amelyik a herékben is expresszálódik, azon a helyen, amelyhez - mint ma már tudjuk - az emberi genom legtöbb változása köthető.
Hogy a herékben kifejeződő gének ezen sajátos változékonyságára fény derüljön persze először szükség volt az említett genom projectek befejezésére. A genomok első körben elvégzett összehasonlítása lényegében megerősítette azt, amit már kisebb DNS régiók összehasonlítása alapján sejthettünk: genetikailag alig különbözünk a csimpánzoktól. Hogy mit értünk pontosan az "alig" alatt, első olvasatban egy picit összezavarhat, mert egyaránt lelhetünk adatot 98.8%-os, illetve 96%-os hasonlóságról az irodalomban. A maga módján egyik szám sem helytelen, csak egy kicsit mást mérnek. Ha igazán objektívek akarunk lenni, akkor a 96% az igazán pontos, mert a 98.8% csak azt méri, hogy a párhuzamba állítható szakaszok között mekkora a hasonlóság. De nem minden szakasznak van meg a párja másik fajban, és ha az ilyen inszerciókat, deléciókat (ún. indeleket) és egyes DNS szakaszok duplikációit is beszámítjuk, akkor a hasonlóság egy kicsit csökken (nota bene, hasonló módon, az említett hatások figyelembevételével, köztünk emberek között is kevesebb, mint 99.9%-os a hasonlóság).
Mindez közérthetőbben annyit tesz, hogy az ember és csimpánz között tízszer több különbség van, mint két átlagos ember között és tízszer kevesebb, mint patkány és egér között. A fehérjék 29%-a tökéletesen azonos, és átlagolva a fehérjéink csak egyetlen aminosavban térnek el egymástól.
Az érem másik oldalát tekintve, mindez azonban úgy is leírható, hogy 35 millió bázispár tér el a két genom között (átlagos hosszuk 3 milliárd bázispár), plusz 5 millió indel. Most akkor ez sok változás vagy kevés? Maradjunk annyiban, hogy mint azt mindennap láthatjuk, elegendő. A változások nagy része olyan DNS szekvenciákba esik, amelyeknek mai tudásunk szerint, nincs se fehérje kódoló, se fehérje szabályozó szerepe: például egy ugráló gén lényegesen aktívabb volt a mi őseinkben, mint a csimpánzokéban, így a mi genomunk sokkal több kópiát hordoz ebből. Ez a kutatók számára egyszerre áldás és átok, mert azt jelenti, hogy csak kevés különbség igazán fontos és lényeges, viszont azokat meg kell valahogy találni...
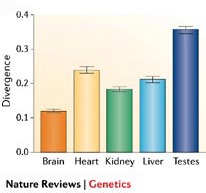 A genom összehasonlítgatások egy másik érdekes hozadéka, hogy a jelek szerint a legváltozékonyabb fehérjéknek egyáltalán nincs köze az agyhoz, ellenben fontos szerepet játszanak az immunitásban és szaporodásban. Mindkét jelenség logikus, és még csak nem is teljesen meglepő, hiszen nagyon hasonló eredményre jutottak az egér-patkány összehasonlítás során is. Immunrendszerünk állandó adaptációra van ítélve a kórokozókkal folytatott "fegyverkezési versenyben", a szaporodásban (különösen spermatogenezisben) fontos gének jó része pedig folyamatosan erős szelekció alatt van, ami meggyorsítja a változásukat. (A spermatogenezisben részt vevő gének egy részének változékonyságára létezhet egyfajta mechanisztikus magyarázat is, mivel sokan a genetikai állományunk legváltozékonyabb kis darabján, az Y kromoszómán található. Y kromoszómából normális esetben sosem fordul elő kettő ugyanabban a személyben, éppen ezért sokkal gyorsabban tud változni, mert az esetleges crossing-overek nem próbálják állandóan homogenizálni a szekvenciáját a populációban. Másrészről azonban ez csak részelges magyarázat lehet a jelenségre, ui. a legkevésbé változó kromoszómán, az X-en levő spermatogenezisben fontos gének is változékonyabbak az átlagnál - ez pedig az aktív szelekció szerepét erősíti.)
A genom összehasonlítgatások egy másik érdekes hozadéka, hogy a jelek szerint a legváltozékonyabb fehérjéknek egyáltalán nincs köze az agyhoz, ellenben fontos szerepet játszanak az immunitásban és szaporodásban. Mindkét jelenség logikus, és még csak nem is teljesen meglepő, hiszen nagyon hasonló eredményre jutottak az egér-patkány összehasonlítás során is. Immunrendszerünk állandó adaptációra van ítélve a kórokozókkal folytatott "fegyverkezési versenyben", a szaporodásban (különösen spermatogenezisben) fontos gének jó része pedig folyamatosan erős szelekció alatt van, ami meggyorsítja a változásukat. (A spermatogenezisben részt vevő gének egy részének változékonyságára létezhet egyfajta mechanisztikus magyarázat is, mivel sokan a genetikai állományunk legváltozékonyabb kis darabján, az Y kromoszómán található. Y kromoszómából normális esetben sosem fordul elő kettő ugyanabban a személyben, éppen ezért sokkal gyorsabban tud változni, mert az esetleges crossing-overek nem próbálják állandóan homogenizálni a szekvenciáját a populációban. Másrészről azonban ez csak részelges magyarázat lehet a jelenségre, ui. a legkevésbé változó kromoszómán, az X-en levő spermatogenezisben fontos gének is változékonyabbak az átlagnál - ez pedig az aktív szelekció szerepét erősíti.)
Ennek ellenére sokakban keltett csalódást, hogy a nyilvánvaló kognitív különbségek ellenére az agyban (is) kifejeződő géneknek mind szekvenciája, mind expressziós szintje kevésbé különbözik, mint más szervekre jellemző géneké (ez látszólag ellent is mondott egy korábbi tanulmánynak, ahol ellenkező eredményre jutottak - mint utólag kiderült, nem azért mert az agyi gének változnak gyorsan, hanem azért, mert azok változtak lassan, amelyekkel összevetették őket). Pedig jobban belegondolva ez sem váratlan: a főemlősök agya mind nagyon bonyolult és szofisztikált szerkezet abszolút értéken mérve, még akkor is, ha a miénkhez mérve egyszerűnek tűnik. Egy ilyen bonyolult szerkezetbe nehéz úgy belenyúlni, hogy ne romoljon el valami, éppen ezért kevesebb mutáció képes "hasznossá tenni magát" és rögzülni.
(Hogy az anthropocentrikusabban gondolkodók se keseredjenek nagyon el, megjegyzendő, hogy bár a jelek szerint a csimpánz-ember szétválás óta az előbbi vonalban összességében több gén változott, mint bennünk, az agyban kifejeződő gének esetében mégis inkább az emberi vonalban következtek be a változások - elsősorban az idegsejtek identitását meghatározó és anyagcseréjét szabályozó fehérjékben.)
De ha a különböző fehérjék, ennyire hasonlítanak, akkor mi a szemmel látható különbség oka? Nos, éppen ez a nagyfokú fehérje azonosság, az ami hirtelen nagyobb hitelt adott egy lassan harminc éves elméletnek, amely Mary-Claire King és Allan Wilson nevéhez fűződik. Ennek értelmében az igazán fontos különbségek, nem a fehérjék szekvenciájában, hanem azok szabályozó-régióiban lesznek (magyarán kifejeződésük helyének és idejének megváltozásában).
És vannak olyan jelek, melyek arra utalnak, hogy tényleg az a helyzet. Amikor olyan DNS szakaszokat hasonlítottak össze (függetlenül azok funkciójától), amelyek viszonylag gyorsan változtak az emberben, de átlagban keveset változtak a gerincesek kialakulása során (ami önmagában arra utal, hogy valami szerepük van, különben szabadon szedhették volna össze a mutációkat), a legtöbb ezek közül nem-(fehérje)kódoló DNS-nek bizonyult. Ezek közül néhány az idegsejtek kapcsolódását segítő fehérjék szabályozó régiója, mások azonban olyan RNS-gének (fehérje nem, csak RNS íródik át róluk), amelyek bizonyos idegsejtekben fejeződnek ki, de funkciójuk ma még ismeretlen.
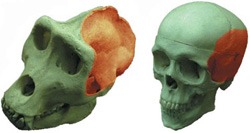 Az előbb említett a változások jelenthetik azt, hogy bizonyos gének egyes kifejeződéséi helyei módosulnak - vagy akár meg is szűnnek. Bizony, bármennyire furcsán hangzik, néhány jellegzetes emberi tulajdonság egyes gének elromlásával hozható összefüggésbe. Ilyen például az aktív szaglóreceptor génjeink számának drámai csökkenése (számos szaglóreceptor mára már csak egy csonk, azaz funkció nélküli pszeudogén formájában lelhető fel a genomunkban. Vagy egy jellegzetes izomfehérje, a MYH16 működésképtelenné válása, amely egyes rágóizmok méretének csökkenését okozta, sokak szerint lehetővé téve, hogy koponyánk (és közvetve az agyunk) nagyobbá válhasson (az izmok kisebbedése tapadásuk helyét is csökkentette, ezáltal téve lehetővé az agykoponya növekedését).
Az előbb említett a változások jelenthetik azt, hogy bizonyos gének egyes kifejeződéséi helyei módosulnak - vagy akár meg is szűnnek. Bizony, bármennyire furcsán hangzik, néhány jellegzetes emberi tulajdonság egyes gének elromlásával hozható összefüggésbe. Ilyen például az aktív szaglóreceptor génjeink számának drámai csökkenése (számos szaglóreceptor mára már csak egy csonk, azaz funkció nélküli pszeudogén formájában lelhető fel a genomunkban. Vagy egy jellegzetes izomfehérje, a MYH16 működésképtelenné válása, amely egyes rágóizmok méretének csökkenését okozta, sokak szerint lehetővé téve, hogy koponyánk (és közvetve az agyunk) nagyobbá válhasson (az izmok kisebbedése tapadásuk helyét is csökkentette, ezáltal téve lehetővé az agykoponya növekedését).
Sok genetikai változás valószínűleg összefügg, de még nem egészen értjük, hogy miképpen. Ilyen összefüggésre utal, hogy sok genetikai betegség esetében (pl. cisztikus fibrózis, fenilketonúria), a hibás fehérje szekvenciája meglepő módon megegyezik az ősi szekvenciával. Ez pedig arra utal, hogy megváltozása összefüggött egész működési kontextusának változásával: azokkal a génekkel/fehérjékkel változott közösen, amelyekkel valamilyen formában funkcionális kapcsolatban van, és mivel a régi szekvencia már nem képes a megváltozott partnerekkel együttműködni, jelenléte betegséget okoz.
Mint látható a puzzle még közel sincs kirakva. De itt tartunk kb. ma. Vannak foltok, ahol már látszik valamilyen mintázat és reméljük, hogy ezek mentén elindulva előbb-utóbb sikerül a teljes történetet kirakni. Nem lesz se rövid, se könnyű, de meg fogja érni, mert nincs kétségem afelől, hogy rengeteg izgalmas összefüggés vár felfedezésre az út során.
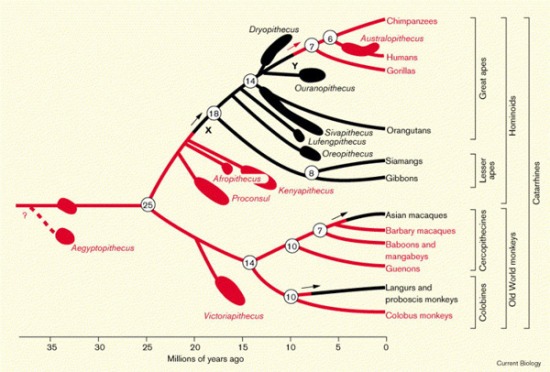
(Az első kép a "The Ape and The Child" project honlapjáról származik.)
Carroll, SB (2003) Genetics and the making of Homo sapiens. Nature 422: 849-857.
Gilbert, SL, Dobyns, WB, Lahn, BT (2005) Genetic links between brain development and brain evolution. Nat Rev Gen 6: 581-590.
Shu, W, Cho, JY, Jiang, Y, Zhang, M, Weisz, D, et al. (2005) Altered ultrasonic vocalization in mice with a disruption in the Foxp2 gene. PNAS 102(27): 9643-9648. doi: 10.1073/pnas.0503739102
Haesler S, Wada K, Nshdejan A, Morrisey EE, Lints T, et al. (2004) FoxP2 expression in avian vocal learners and non-learners. J Neurosci 24: 3164–3175.
Enard, W, Przeworski, M, Fisher, SE, Lai, CSL, Wiebe1, V, Kitano, T, Monaco, AP, Pääbo, S (2002) Molecular evolution of FOXP2, a gene involved in speech and language. Nature 418: 869-872.
Lai, CSL, Fisher, SE, Hurst, JA, Vargha-Khadem, F, Monaco, AP (2001) A forkhead-domain gene is mutated in a severe speech and language disorder. Nature 413: 519−523.
Li, WS, Saunders, MA (2005) The chimpanzee and us. Nature 437:50-51.
Khaitovich, P, Enard, W, Lachmann, M, Pääbo, S (2006) Evolution of primate gene expression. Nat Rev Gen 7: 693-702.
Khaitovich, P, Hellmann, I, Enard, W, Nowick, K, Leinweber, M, et al. (2006) Parallel patterns of evolution in the genomes and transcriptomes of humans and chimpanzees. Science 309: 1850-1854.
Uddin, M, Wildman, DE, Liu, G, Xu, W, Johnson, RM, et al. (2004) Sister grouping of chimpanzees and humans as revealed by genome-wide phylogenetic analysis of brain expression profiles. PNAS 101:2957-2962.
Wang, HY, Chien, HC, Osada, N, Hashimoto K, Sugano, S, et al. (2007) Rate of evolution in brain-expressed genes in humans and other primates. PLoS Biol 5(2): e13.
Dorus, S, Vallender, EJ, Evans, PD, Anderson, JR, Gilbert, SL, et al. (2004) Accelerated evolution of nervous system genes in the origin of Homo sapiens. Cell 119: 1027-1040.
Fu N, Drinnenberg I, Kelso J, Wu J, Pääbo S, et al. (2007) Comparison of Protein and mRNA Expression Evolution in Humans and Chimpanzees. PLoS ONE 2(2): e216. doi:10.1371/journal.pone.0000216
Bakewell, MA, Shi, P, Zhang, J (2007) More genes underwent positive selection in chimpanzee evolution than in human evolution. PNAS 104: 7489-7494.
King, MC, Wilson, AC (1975) Evolution at two levels in humans and chimpanzee. Science 188: 107-116.
Prabhakar, S, Noonan, JP, Pääbo, S, Rubin, EM (2006) Accelerated evolution of conserved noncoding sequences in humans. Science 314 786.
Pollard, KS, Salama, SR, King, B, Kern, AD, Dreszer, T, et al. (2006) Forces shaping the fastest evolving regions in the human genome. PLoS Genet 2(10): e168. DOI: 10.1371/journal.pgen.0020168
Pollard, KS, Salama, SR, Lambert, N, Lambot, MA, Coppens, S, et al. (2006) An RNA gene expressed during cortical development evolved rapidly in humans. Nature 443: 167-172.
Stedman, HH, Kozyak, BW, Nelson, A, Thesier, DM, Su, LT, et al. (2004) Myosin gene mutation correlates with anatomical changes in the human lineage. Nature 428: 415-418.
Gilad, Y, Man, O, Pääbo S, Lancet, D (2003) Human specific loss of olfactory receptor genes. PNAS 100: 3324-3327.
Gilad, Y, Wiebe, V, Przeworski, M, Lancet, D, Pääbo, S (2004) Loss of Olfactory Receptor Genes Coincides with the Acquisition of Full Trichromatic Vision in Primates. PLoS Biol 2(1): e5 doi:10.1371/journal.pbio.0020005
Rhesus Macaque Genome Sequencing and Analysing Consortium Evolutionary and Biomedical Insights from the Rhesus Macaque Genome. Science 316 222-234.

Utolsó kommentek