(Vendégmunkásunk, SexComb tollából.) Újabb ÉRTEM cikk kerül a
boncasztalra, ezúttal a molekuláris genetika tárgyköréből, Biokémiai
hasonlóságok címmel. Az eredeti itt található.
"A biokémiai taxonómia
egyik legfontosabb eljárása a fehérjén belüli aminosav-sorrend (szekvencia) meghatározása,
illetve a DNS-ben lévő bázishármasok (tripletek) sorrendjének megállapítása.
A kutatók DNS- és fehérje-szekvencia analizátorokat alkalmaznak
e sorrendek a meghatározásához."
 A molekuláris biológia fejlődésével
lehetővé vált gyorsan, olcsón meghatározni rövidebb DNS szakaszok
bázissorrendjét, ugyanis a DNS szekvenálás gépesíthető, egyetlen
közepesen képzett munkatárs képes kiszolgálni egy szekvenáló-gépet,
így egyetlen reakció költsége öt-tíz euró körül mozog, amely
körülbelül ezer-ezerötszáz bázis leolvasását teszi lehetővé.
A fehérjék aminosavsorrendjének meghatározására is van lehetőség,
ám ezek az eljárások lassúak így meglehetősen költségigényesek,
ilyen célokra szinte sohasem használják őket. Az adatbázisokban
szereplő fehérjeszekvenciák túlnyomó többsége egyszerűen a DNS
bázissorrendje alapján "lefordított" fehérje. A DNS esetében
bázishármasokról csak a fehérjéket kódoló szakaszok esetében
beszélhetünk, mivel valóban három bázis határoz megy egy aminosavat,
ám a fehérjéket nem kódoló szakaszok esetében ez a szóhasználat
értelmezhetetlen, így innentől csak DNS bázissorrendről beszélnék,
a cikk szerzője is nyilván erre gondolt. A DNS bázissorrend vizsgálatának
óriási előnye, hogy matematikai módszerekkel is elemezhető, így
nem jelenik meg az a pontatlanság, amelyet az egyes külsődleges jellegek
értékelése szükségképpen tartalmaz, hiszen nem önkényesen kijelölt
jellegeket vizsgálunk. A cikk szerzőjének választása éppen a citokróm
c fehérjére esett, maradjunk hát ennél a példánál. A citokróm
c a mitokondriumban található, a sejt energiatermelési folyamatai
során egy elektront szállít. Éppen mivel ilyen alapvető folyamatban
vesz részt, kevéssé változékony fehérje, homológja könnyen azonosítható
az éppen vizsgált szervezetben, ráadásul a hossza kevéssé változékony,
így az egyes fehérjék könnyen összehasonlíthatóak, éppen ezért
alkalmas evolúciós vizsgálatokra.
A molekuláris biológia fejlődésével
lehetővé vált gyorsan, olcsón meghatározni rövidebb DNS szakaszok
bázissorrendjét, ugyanis a DNS szekvenálás gépesíthető, egyetlen
közepesen képzett munkatárs képes kiszolgálni egy szekvenáló-gépet,
így egyetlen reakció költsége öt-tíz euró körül mozog, amely
körülbelül ezer-ezerötszáz bázis leolvasását teszi lehetővé.
A fehérjék aminosavsorrendjének meghatározására is van lehetőség,
ám ezek az eljárások lassúak így meglehetősen költségigényesek,
ilyen célokra szinte sohasem használják őket. Az adatbázisokban
szereplő fehérjeszekvenciák túlnyomó többsége egyszerűen a DNS
bázissorrendje alapján "lefordított" fehérje. A DNS esetében
bázishármasokról csak a fehérjéket kódoló szakaszok esetében
beszélhetünk, mivel valóban három bázis határoz megy egy aminosavat,
ám a fehérjéket nem kódoló szakaszok esetében ez a szóhasználat
értelmezhetetlen, így innentől csak DNS bázissorrendről beszélnék,
a cikk szerzője is nyilván erre gondolt. A DNS bázissorrend vizsgálatának
óriási előnye, hogy matematikai módszerekkel is elemezhető, így
nem jelenik meg az a pontatlanság, amelyet az egyes külsődleges jellegek
értékelése szükségképpen tartalmaz, hiszen nem önkényesen kijelölt
jellegeket vizsgálunk. A cikk szerzőjének választása éppen a citokróm
c fehérjére esett, maradjunk hát ennél a példánál. A citokróm
c a mitokondriumban található, a sejt energiatermelési folyamatai
során egy elektront szállít. Éppen mivel ilyen alapvető folyamatban
vesz részt, kevéssé változékony fehérje, homológja könnyen azonosítható
az éppen vizsgált szervezetben, ráadásul a hossza kevéssé változékony,
így az egyes fehérjék könnyen összehasonlíthatóak, éppen ezért
alkalmas evolúciós vizsgálatokra.
"Most nézzük meg
a selyemhernyó lepkéjének sorát (a táblázat 15. oszlopa),
és haladjunk lefelé
a gerincesek között osztályról osztályra. Figyeljük meg, hogy
e rovar citokróm c fehérjéje közel ugyanolyan mértékben különbözik
olyan egymástól igen eltérő szervezetekétől, mint az ember, a
pingvin, a teknős, a tonhal és az orsóhal. Figyelembe véve az említett
szervezetek közti óriási változatosságot, meglepő, hogy citokróm
c fehérjéjük tekintetében mindannyian majdnem ugyanolyan százalékban
különböznek a selyemhernyó lepkéjétől."
 Ez miért lenne meglepő? Bátorkodtam
az ábrán színekkel jelölni az egyes élőlénycsoportokat: Pirosak
az emlősök és az egy szem erszényes, a kenguru, kékek a gerincesek.
Zöld az egyetlen ízeltlábú, sárga az egyetlen növény, narancssárga
az egyetlen baktérium. A lepke és az ember utolsó közös őse ugyanaz
az állat volt, mint a lepke és az orsóhal utolsó közös őse, hiszen
ezek az élőlények mind gerincesek, amelyek teljesen különálló
törzset alkotnak. Egy ló rendszertani
értelemben pont annyira különbözik egy lepkétől, mint egy
ponty. A táblázat adatai éppen ezt támasztják alá. Mindez miért
mond ellent az evolúciónak?
Ez miért lenne meglepő? Bátorkodtam
az ábrán színekkel jelölni az egyes élőlénycsoportokat: Pirosak
az emlősök és az egy szem erszényes, a kenguru, kékek a gerincesek.
Zöld az egyetlen ízeltlábú, sárga az egyetlen növény, narancssárga
az egyetlen baktérium. A lepke és az ember utolsó közös őse ugyanaz
az állat volt, mint a lepke és az orsóhal utolsó közös őse, hiszen
ezek az élőlények mind gerincesek, amelyek teljesen különálló
törzset alkotnak. Egy ló rendszertani
értelemben pont annyira különbözik egy lepkétől, mint egy
ponty. A táblázat adatai éppen ezt támasztják alá. Mindez miért
mond ellent az evolúciónak?
"Ez a felfedezés azért olyan megdöbbentő, mert ellentmond a darwini elvárásoknak. Az evolúciós skálán a selyemhernyó
lepkéjétől felfelé haladva azt várnánk, hogy molekuláris szinten egyre nagyobb különbségeket
tapasztalunk majd."
"A darwinizmusból
az következne, hogy ha összehasonlítjuk a ma
élő szervezeteket, akkor nagyobb molekuláris távolságot tapasztalunk
a rovaroktól a kétéltűekig, mint a ma
élő halakig, még nagyobb távolságot a hüllőkig,
és még ennél is nagyobbat az emlősökig. A valóság azonban nem
ezt a mintát igazolja vissza."
Miért várnánk nagyobb különbségeket?
Ezelőtt körülbelül ötszáz–ötszázötven millió éve élt egy
jószág, amelynek távoli leszármazottainak egyik csoportjából lettek
az ízeltlábúak, másik csoportjából a gerincesek. Miután különvált
ez a két leszármazási vonal, nyilván nem hoztak létre közös utódokat,
véletlenszerű mutációk történtek bennük. A két vonal szétválása
után mindkét leszármazási vonalban mutációk történtek a citokróm
c génben, ám ezek egymástól független események kellettek,
hogy legyenek, mivel nem tudunk róla, hogy ízeltlábúak valaha kereszteződtek
volna gerincesekkel. A cikk felvetése teljesen téves, nem igazán
értem, milyen gondolatmenet alapján jutott a szerző erre a következtetésre.
A ma élő selyemlepke nem őse az embernek. Azt hiszem, ennek a belátásához
egy kis józan észen kívül semmi más nem szükséges. A két
élőlénynek a távoli múltban valóban volt egy közös
őse, ám azóta egymásról mit sem tudva változtak, teljesen külön
utakon.
"Például a darwinisták
a kétéltű kecskebékát egy többszörös elágazás termékének
tekintik, amely a kétéltűek hüllőkhöz vezető
ágáról való leágazás után ment végbe. Ez azt jelenti, hogy bár
a kecskebéka a gerincesek egy olyan rendjének a tagja, amely a hüllőknek
is őse, ő maga nem őse a hüllőknek. "
"A darwinizmusból
az következne, hogy ha összehasonlítjuk a ma
élő szervezeteket, akkor nagyobb molekuláris távolságot tapasztalunk
a rovaroktól a kétéltűekig, mint a ma
élő halakig, még nagyobb távolságot a hüllőkig,
és még ennél is nagyobbat az emlősökig."
"A hagyományos darwini
forgatókönyv szerint a kétéltűek a halak
és a szárazföldi gerinces állatok közötti
átmenetek."
Itt egy alapvető
tévedéssel van dolgunk: A ma élő
kecskebéka nem őse a hüllőknek. Azért nem, mert hüllők már
jó néhány millió éve élnek a földön, így ha a tegnap befogott
kecskebéka az ő elődjük, ez azt feltételezné, hogy a nyomorult
jószág több tízmillió éves matuzsálem, hiszen ő nemzette azokat
az utódokat, akikből később a hüllők alakultak ki. Maradjunk annyiban,
hogy ez kizárható. Annak idején létezett egy állatcsoport, amely
akkoriban a mai kétéltűekre hasonlított inkább, őbelőlük alakultak
ki idővel a mai hüllők és a mai kétéltűek. A ma élő béka nyilvánvalóan
abba a csoportba tartozik, amely nem indult el a hüllővé válás
útján, hanem megőrizte ennek az ősi csoportnak a jellegzetességeit.
Ez nem jelenti azt, hogy a hozzá vezető leszármazási vonalban nem
történtek mutációk. Nyilván ebben a leszármazási vonalban is
számos mutáción átesett már a citokróm c gén, mert a hüllők
kialakulása óta eltelt időben a kétéltűek is változtak. Az
érdekes az, hogy mindezzel az első
idézet tanúsága szerint a szerző
is tökéletesen tisztában van, azonban valamiért mégis egy ennek
ellentmondó gondolatmenetet követ az egész cikk során,
így tudatosan egy sohasem létezett
elmélet cáfolatát fejti ki. A kérdés csak az, hogy mindezt miért?
Ugyanez a helyzet a hallal is. A ma élő ponty nem lehet a ma élő
ló őse, hacsak nem feltételezzük, hogy a lerakott ikráiból csikók
kelnek ki. Annak idején élt egy hal, a két csoport utolsó közös
őse, amelynek utódai meghódították a szárazföldet, belőlük
fejlődtek ki a szárazföldi gerincesek. Ugyanezen hal leszármazottainak
egy másik csoportja nem indult el ezen az úton, maradt, ahol volt,
a vízben. Egy hal és egy ló utolsó közös őse pontosan ugyanaz
az egyed volt, mint egy hal és egy teknős közös őse. Miért kellene
egy hal citokróm c fehérjéjének jobban hasonlítani egy teknősére,
mint egy lóéra? A lóhoz és a teknőshöz vezető leszármazási
vonal pontosan ugyanabban az időpillanatban vált el a halhoz vezető
leszármazási vonaltól. Miért kellene köztük nagymértékű különbséget
látni? Rajzoltam egy ábrát a boncolgatott cikk által felvetett kérdésről.
A citokróm c fehérje egy rövid szakaszának az elképzelt változásait
tüntettem fel rajta.

Ez a kitalált modell mutatja
be a cikk hibás alapvetését, vagyis, hogy a gerincesek citokróm
c fehérjéjének egyre jobban kellene különböznie a selyemlepke
azonos fehérjéjétől. A képlet egszerű: Selyemlepke-Ponty: egy
eltérés. Selyemlepke-Béka: két eltérés. Selyemlepke-Teknős: három
eltérés és így tovább. Az adatok valóban nem ezt mutatják. Mi
hát a hiba?
Az első
hiba a rosszul megválasztott vonatkoztatási pont. A szerző valamiért
kiindulási alapként a ma élő selyemlepke citokróm c fehérjéjét
kezeli őstípusként, amelyből a többi kifejlődött. A két élőlénycsoportnak
valóban közös őse volt, ám ez az állat ötszázmillió évnél
is régebben élt. Ha a mai selyemlepke citokróm c fehérjéjét választjuk
viszonyítási alapnak, akkor ezzel feltételezzük, hogy az azonos
az ötszázmillió éve élt közös ős citokróm c fehérjéjével.
Felmerül a kérdés, hogy ez lehetséges –e? A Uniprotadatbázisból bátorkodtam kikeresni
néhány ízeltlábú állat citokróm c fehérjéjét és összehasonlítani
ezeket. Az eredmények az alábbi táblázatban láthatóak:
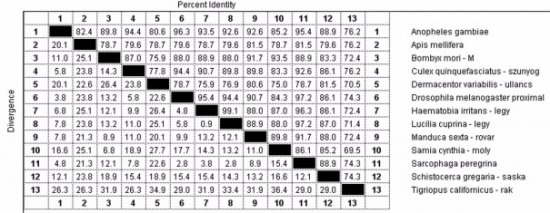
Minden vizsgált szervezet
az ízeltlábúak törzsébe tartozik, azonban látható, hogy az ő
citokróm c fehérjéik is nagymértékben különböznek egymástól.
Például a mézelő méh (Apis mellifera) citokróm c fehérjéje csak
78,7 százalékban egyezik meg a selyemlepke (Bombyx mori) citokróm
c fehérjéjével, ami elég nagy különbség. Be kell, hogy lássuk,
hogy az ízeltlábúak törzsében is történtek mutációk az utóbbi
ötszázmillió évben, az ő citokróm c fehérjéik is különböznek
egymástól. (Az lenne a furcsa, ha nem történtek volna.) Ezek
után, ha valaki abból indul ki, hogy a ma
élő selyemlepke citokróm c fehérjéje ugyanaz a fehérje, amelyből
annak idején a gerincesek citokróm c fehérjéi kialakultak, akkor
legalábbis illene bizonyítania, hogy miért
éppen az ezt a fehérjét kódoló
gén kerülte el a mutációkat az utóbbi
ötszázmillió évben? Miért olyan különleges ez a mai selyemlepkéhez
vezető leszármazási vonal, hogy változatlanul megőrizte az ősi
citokróm c fehérjét, miközben a többi ízeltlábúban rengeteg
mutáció történt e fehérjében? Miért nem a mézelő méh, vagy
a maláriaszúnyog citokróm c fehérjéjét választotta kiindulási
alapnak a szerző? Ugyanez a hiba a ponty vonatkoztatási pontként
használatával is, feltételezhetően az ő citokróm c fehérjéje
sem egyezik már meg az emlősökkel közös ős hasonló fehérjéjével.
A második hiba az, hogy
elfeledkezik róla, hogy ezek az élőlények az utóbbi néhány százmillió
évben is éltek, szaporodtak, nyilván mutációk is bekövetkeztek
bennük. Amint elválik egymástól két leszármazási vonal, onnantól
mindkét vonalban véletlenszerűen, ráadásul egymástól teljesen
függetlenül történnek mutációk. Ha ezt is számításba vesszük,
nyugodtan átrajzolhatjuk a fenti egyszerű ábránkat. Ha feltételezzük,
hogy egyetlen élőlény sem ellenálló a mutációkkal szemben –
mint ahogy minden kísérleti adat ezt bizonyítja – és minden egyes
lépésben egy újabb véletlen mutációt illesztünk a már meglévő
fehérjékbe, azonnal átalakul a kép:

Ha ugyanígy megszámoljuk
az eltéréseket, alapvetően más eredményre jutunk: Selyemlepke-Ponty:
hét eltérés. Selyemlepke-Béka: nyolc eltérés. Selyemlepke-Teknős:
nyolc eltérés. Selyemlepke-Ló: nyolc eltérés. Selyemlepke-Ember:
nyolc eltérés. Ponty-Béka: négy eltérés. Ponty-Teknős: négy
eltérés. Ponty-Ló: négy eltérés. Ponty-Ember: öt eltérés. Mivel
véletlenszerűen illesztettem be a változásokat egy dobókocka segítségével,
így némileg ingadozik az eredmény, ám az összkép nem változik:
Körülbelül azonos eredményt kapunk vissza az egyes leszármazási
vonalakban, a folyamat végére előálló selyemlepke-fehérje éppen
úgy különbözik az ember és a béka fehérjétől, eltűnik az előző,
hibás modell alapján várt különbség. Ha megnézzük a valós
eredményeket, éppen ugyanezt látjuk. Úgy tűnik, megint sikerült
magyarázatot találnunk egy természeti jelenségre az értelmes tervező
mellőzésével is: A selyemlepke ősei semmilyen módon sem keveredtek
a gerincesek őseivel vagy ötszázmillió éve, így a két csoportban
egymástól független mutációs események történtek. Ráadásul
ugyanígy a kétéltűek sem váltak hirtelen ellenállóvá a mutációkkal
szemben csak azért, mert kialakultak a hüllők, majd az emlősök.
"Először is: a tudomány
szerint a mutációk aránya a generációs
időhöz viszonyul, s a különböző
molekulák mutációs aránya azonos az egyes generációkban.
"
Ez nem tudom, melyik tudomány
szerint van így, a biológia tudománya például semmi ilyesmit sem
állít. Lássunk egy példát! Gondolom mindenki hallott már arról,
hogy minél idősebbek a szülők, annál nagyobb eséllyel szenved
a születendő gyermekük valamilyen örökletes betegségben. Vegyük
például a már emlegetett Down-kórt. Ismert, hogy a negyven felett
szülő nők gyermekiben sokkal gyakoribb a Down-kór, mint a fiatalon
szülő nők esetében. Ha elolvassuk a Wikipedia idevágó fejezetét, még egy ábrát
is találunk:
 Vagyis a harminc évesen szülő
nőknek egy ezrelék eséllyel születik Down-kóros gyermeke, míg
a negyven évesen szülő nőknek egy százalék esélye van Downos
utód létrehozására. A nők testében a petesejtek a magzati élet
során kialakulnak, soha többet nem osztódnak. Ha egy nő harminc
évesen szül, pontosan ugyanakkor keletkezett petesejtje termékenyül
meg, mint ha negyven évesen szülne. Akkor mégis miért különbözik
a cikk írója szerint az öröklött betegségek, ez esetben a Down-kór
előfordulásának aránya, ha szerinte nemzedékenként azonos mennyiségű
mutáció történik? A vizsgált esetben pontosan egy nemzedék választja
el az anyát gyermekétől, mégis a mutáció esélye éppen tízszeres
ha negyven évesen szül egy nő, mint ha harminc évesen alapít családot.
Hogyan lehet ez? Nagyon egyszerű, a
mutációk gyakorisága nem azonos nemzedékenként, függ a nemzedékek
hosszától is, mivel egy ivarsejtben osztódás nélkül, nyugalomban
is történhetnek mutációk. Ezzel azt hiszem sikerült is cáfolni
az idézett állítást.
Vagyis a harminc évesen szülő
nőknek egy ezrelék eséllyel születik Down-kóros gyermeke, míg
a negyven évesen szülő nőknek egy százalék esélye van Downos
utód létrehozására. A nők testében a petesejtek a magzati élet
során kialakulnak, soha többet nem osztódnak. Ha egy nő harminc
évesen szül, pontosan ugyanakkor keletkezett petesejtje termékenyül
meg, mint ha negyven évesen szülne. Akkor mégis miért különbözik
a cikk írója szerint az öröklött betegségek, ez esetben a Down-kór
előfordulásának aránya, ha szerinte nemzedékenként azonos mennyiségű
mutáció történik? A vizsgált esetben pontosan egy nemzedék választja
el az anyát gyermekétől, mégis a mutáció esélye éppen tízszeres
ha negyven évesen szül egy nő, mint ha harminc évesen alapít családot.
Hogyan lehet ez? Nagyon egyszerű, a
mutációk gyakorisága nem azonos nemzedékenként, függ a nemzedékek
hosszától is, mivel egy ivarsejtben osztódás nélkül, nyugalomban
is történhetnek mutációk. Ezzel azt hiszem sikerült is cáfolni
az idézett állítást.
"Ezen kívül az egyes
fehérjék mutációs aránya még ugyanazon a fajon belül is eltérő.
Ez azt jelenti, hogy ahhoz, hogy
a molekuláris óra elmélete helyes legyen, nem egy, hanem több ezer molekuláris
órával kellene számolni."
A "molekuláris
óra" nem létezik, egyszerűen egy jelenség leírása, nem valós,
működő szerkezet. Egy "molekuláris óra" egy homológ
szakasz két vagy több szervezet genomjában, amelynek a különbségeit
vizsgáljuk. Egy ilyen szakaszt, azaz egy "molekuláris órát"
a kísérletet végző kutató jelöl ki, az adott kísérlet szempontjai
alapján. Mutációk véletlenszerűen történnek, tehát nagyon nagy
időszakokat nézve egy adott DNS szakaszban körülbelül adott mennyiségű
mutáció keletkezik. Mindez annyit jelent, hogy két különböző
élőlényben ugyanaz a homológ, azaz egymásnak megfelelő DNS szakasz
minél több mutációt tartalmaz, annál régebben váltak külön,
azaz annál régebben élt a két élőlény utolsó közös őse. A
fenti táblázatból is mindössze annyi olvasható le, hogy az ember
és a majom utolsó közös őse sokkal később élt, mint az ember
és a béka, vagy az ember és a molylepke utolsó közös őse. Egyetlen
élőlényben sem ketyeg a szerző által feltételezett "molekuláris
óra", hiszen semmi szükségük sem lenne rá, mondjuk egy békának
mi szüksége lenne rá, hogy számon tartsa, mikor élt a halakkal
közös őse? Meg kell, hogy állapítsuk, a szerzőnek fogalma sincs
arról, mit is nevezünk ebben az esetben "molekuláris órának".
Az adott genomban lévő "molekuláris órák" száma így
egyedül a szemlélőtől függ, azaz annyi molekuláris óra létezik
egy genomban, amennyit kijelölünk. Ha a citokróm c gént vizsgáljuk,
akkor egy molekuláris óránk van, ha mellé vesszük az aktint
és az RNS polimeráz II gént, akkor máris három. Amennyiben
megfelelő mennyiségű időnk és pénzünk van, több ezer molekuláris
órával is számolhatunk, ám ennek nem sok értelme lenne. Ha megbízható
törzsfát akarunk felállítani, érdemes több gén alapján dolgozni,
mivel véletlen események átlagát mérjük egy ilyen vizsgálattal,
így az egyes gének vizsgálatával kapott eredmények kisebb-nagyobb
mértékben eltérhetnek egymástól, ám túlzásokba sem érdemes
esni.
" Az azonos univerzumon
belüli működés szükségessége az
összes élő szervezettel szemben azonos fizikai
és kémiai igényeket támaszt. Egy intelligens cselekvő
számára logikus és egyszersmind hatékony is az
élőlényeket közös biokémiai alapon megtervezni."
Számomra éppen ez az értelmes
tervezés elméletének leghihetetlenebb része. Ha már úgyis ugyanazt
a folyamatot végzi egy fehérje, nevezetesen a citokróm c egy elektront
szállít a mitokondriumban, akkor minek belőle ennyi különböző
változat? Ha a fenti állítás igaz, azaz azonos feltételekre az
értelmes tervező azonos molekulákat tervez, miért különböznek
mégis az egyes élőlényekben leírt citokróm c fehérjék? Hogy
az eredeti cikk példájánál maradjak: Nagyon nehéz elképzelni,
hogy az ember és a rhesus majom anyagcseréje annyira különbözne,
hogy új citokróm c fehérjét kellett hozzá tervezni. Akkor meg minek?
Nem gondolnám, hogy az ember és a ló szervezete olyan alapvető folyamatokban
különbözne, hogy az ő citokróm c fehérjéjük már tizenkét
százalékban kell hogy különbözzön. Miért vette mégis a fáradtságot
az értelmes tervező, hogy ennyi különböző citokróm c fehérjét
tervezzen, ha úgyis "azonos fizikai és kémiai igények"
–nek kell megfelelnie? Ennek fényében vegyük szemügyre az ÉRTEM
mozgalom egy régebbi cikkét a fehérjékről (Molekuláris gépezetek):
" Viszont, mivel a
csilló összetettsége egyszerűsíthetetlen, ezért nem lehetnek működőképes
elődei. Miután az egyszerűsíthetetlenül
összetett csillónak nem lehet működő
őse, ezért nem jöhetett létre természetes kiválasztódás révén,
amely igényli a működés folytonosságát a munkájához. A természetes
kiválasztódás tehetetlen, ha nincs működés, melyet kiválaszthatna."
" Az egyszerűsíthetetlen
összetettségre tengernyi további példa van, többek között a fehérjeszállítás,
a véralvadás, a zárt körkörös DNS, az elektronszállítás, a
baktérium ostora, a telomerek, a fotoszintézis, annak szabályozása,
hogy hogyan készüljön az RNS-másolat a génről,
és még számos hasonló."
"Ahogy a
képcsövet, a vezetékeket, az anyacsavarokat
és a fémcsavarokat tartalmazza a tévékészülék,
úgy számos fehérje olyan struktúráknak az alkotórésze, amelyek
csak akkor működnek, hogyha valóban minden alkotórész a helyén
van."
Tehát tulajdonképpen az elektronszállítás,
amelyet a mitokondriumban éppen a citokróm c végez egy egyszerűsíthetetlenül
összetett rendszer. Az ember és a búza elektronszállítást végző
fehérjéje majdnem negyven százalékban különbözik egymástól.
Az ember és a Rhodospirillum rubrum baktérium elektronszállítást
végző fehérjéje hatvanhat százalékban különbözik egymástól.
Ha egy egyszerűsíthetetlenül összetett rendszer egyik elemének
a kétharmada megváltozhat, és zavartalanul működik az nem üti
agyon eleve az egyszerűsíthetetlen összetettség tényét? Ha a "molekuláris
gépezet" kétharmada nyugodtan kicserélődhet, akkor nem lehetséges,
hogy mégis csak nyílik némi tere a véletlen változásoknak is?
Ha egy egyszerűsíthetetlenül összetett rendszer egyik alkatrészéből
több ezer, egymástól nagy mértékben, akár kétharmadában is
különböző változat létezik, akkor tulajdonképpen miért is egyszerűsíthetetlen?
Olvassuk csak el az "egyszerűsíthetetlenül összetett" meghatározását:
"Ebben Behe azokat
a rendszereket nevezi egyszerűsíthetetlenül komplexnek (EK), amelyek:
„számos jól illeszkedő, egymással kölcsönhatásban
álló részből állnak, melyek hozzájárulnak az alapvető funkcióhoz,
s ahol e részek bármelyikének eltávolítása a rendszer működésének
megszűnéséhez vezet”. "
Gondolkodjunk el ezen egy pillanatra!
"számos jól illeszkedő, egymással kölcsönhatásban
álló részből állnak" "csak akkor működnek, hogyha valóban
minden alkotórész a helyén van"
Vagyis akkor az egyik elem módosítása mindenképpen feltételezi
a rendszer összes többi elemének módosítását is. Ha nem így
lenne, akkor bizony azonnal tere nyílna az evolúciónak is, hiszen
ha az egyes "alkatrészek" a többitől függetlenül is nagymértékben
megváltozhatnak, akkor már nem is olyan hihetetlen a feltételezés,
hogy bizony véletlen folyamatok is kialakíthatták a rendszert alapelemeiből.
Tulajdonképpen ezek szerint annyi egyszerűsíthetetlenül összetett
elektronszállító rendszer létezik, ahány citokróm c fehérje.
Ez mennyi lehet? Százezer? Mondjuk egymillió? Nagyon kevés élőlény
genomját ismerjük, de így is rengeteg különböző citokróm c fehérjét
találtunk eddig, amelyek egymástól nagyon nagy mértékben különböznek.
Ez bizony ugyanannyi új, egyszerűsíthetetlenül összetett rendszert
jelent, amelyet bizony az értelmes tervezőnek egyesével újra meg
kellett terveznie, hiszen nem alakulhatott ki valamiféle egyszerűbb
rendszerből, mindet újra fel kellett találnia, mégpedig saját magának.
Nem tűnik ez kicsit sok felesleges munkának? "Az azonos univerzumon
belüli működés szükségessége az
összes élő szervezettel szemben azonos fizikai
és kémiai igényeket támaszt." Mégis zavarbaejtően sok
egyszerűsíthetetlenül összetett rendszer létezik egy egyszerű,
jól körülhatárolható feladatra is, nevezetesen az elektronszállításra.
"Egy intelligens cselekvő számára logikus
és egyszersmind hatékony is az élőlényeket közös biokémiai alapon
megtervezni." Valóban. Viszont felmerül a kérdés, hogy
miért nem így cselekedett akkor? Verejtékes munkával újra és újra
megalkotott ugyanarra a feladatra akárhány, egymástól nagymértékben
különböző egyszerűsíthetetlenül összetett rendszert. Miért?
A szokásos érvelés: "Mért tervezték meg az Oldsmobilt,
amikor egy Mazda, vagy egy Ferrary sokkal jobb." (ID_EGEN hozzászólása egy korábbi cikkhez). No de miért
hagyja az értelmes tervező szanaszét heverni a régebbi változatokat,
miért nem a jobbat használja mindenhol? Ha választhatnék, hogy egy
Oldsmobillal induljak el valahová, vagy egy Ferrarival, én bizony
az utóbbi mellett döntenék. Azonkívül melyik az Oldsmobil és melyik
a Mazda, melyik a Ferrari? A ló citokróm c fehérjéje a legújabb,
tökéletes változat, vagy a lepkéé? Netán a rizsé? Az emberé?
De akkor miért van a többi? Ezek az élőlények is élnek, láthatóan
nagyon is jól működik a citokróm c fehérjéjük, legalábbis nem
tudok róla, hogy az ember/ló/lepke elektronszállító rendszeréről
valaha leírták volna, hogy nagyságrendekkel rosszabbul működik,
mint a többieké. Így sajnos visszajutottunk az alapkérdéshez, vagyis,
hogy az értelmes tervező miért tervez ugyanarra a feladatra sok különböző
rendszert, ha "...logikus és egyszersmind hatékony is az
élőlényeket közös biokémiai alapon megtervezni." Mi erre
az értelmes tervezés mozgalom magyarázata?
Ha már itt tartunk, vegyünk
szemügyre egy másik példát, mondjuk teljesen véletlenül az állandó
vesszőparipámat, a Drosophila melanogastert, azaz az ecetmuslicát.
Ha kikeressük a citokróm c fehérjét a Drosophila genom adatbázisból,
azt a meglepő tényt tapasztaljuk, hogy a légynek nem egy, hanem két
citokróm c génje van.
Ezek egymás mellett helyezkednek el a második kromoszómán, citokróm
c proximal és citokróm c distal néven találhatóak meg a Flybase
oldalon.

Ha összehasonlítjuk a két
gén által kódolt fehérjék szerkezetét, újabb meglepetés vár
ránk: A citokróm c proximal fehérje száznyolc aminosav méretű,
míg a citokróm c distal százöt aminosav hosszúságú. Ha egy, a
fentihez hasonló illesztést végzünk egy megfelelő programmal, azt
tapasztaljuk, hogy a két fehérje harminc százalékban különbözik
egymástól.

Ez a különbség a fenti táblázaton
a selyemlepke-ember távolságnak felel meg. Ezt hogyan magyarázza
az értelmes tervezés elmélete? Adott egy egyszerűsíthetetlenül
összetett rendszer, aminek egy alkatrészéből egyetlen egyeden belül
is két változat létezik, amelyek ráadásul egymástól is gyökeresen
különböznek. Miért tervezett az értelmes tervező ebbe az egy jószágba
két citokróm c fehérjét? Itt bizony nem áll az a magyarázat, hogy
különböző körülményekhez kellett alkalmaznia az egyes változatokat.
De akkor hogyan lehet egy egyszerűsíthetetlen rendszerből ugyanabban
az egyedben kettő? Nyilván az egyik jobban működik, mint a másik,
de akkor minek van ott mellette a rosszabbik is? Ugye a magyarázat
erre is készen áll: "Mért tervezték meg az Oldsmobilt, amikor
egy Mazda, vagy egy Ferrary sokkal jobb."
Na jó, de akkor minek építette be az értelmes tervező ezt a két
rendszert ugyanabba az állatba?
Olvassuk el, mit is csinál ez a két fehérje
a légyben! A citokróm c fehérje az elektronszállítás mellett
az apoptózis folyamatában is szerepet játszik. A cyt-c-d
gén csak a hím csíravonalban fejeződik ki, míg a cyt-c-p
az egész szervezetben. A cyt-c-d
gén elrontása csak a spermiumképzést gátolja, míg a cyt-c-p
gén elrontása az embrió korai halálához vezet. Arama E. és munkatársai
2006 –ban elvégeztek egy kísérletet: Elrontották a cyt-c-d
gént, e mutáció hatására zavar keletkezett a spermiumképzésben,
a hímek terméketlenek lettek, ám ha ezekben a sejtekben mesterségesen
kifejeztették a cyt-c-p vagy a cyt-c-d
gént, megállapították, hogy mindkét fehérje képes ellátni a
citokróm c distal fehérje feladatát, mindkét csoport hímjei termékenyek
voltak. Ezek után kipróbálták, mi történik, ha a cyt-c-p
gént rontják el: Mint ahogy már írtam, ezek az állatok a petéből
sem kelnek ki, nagyon korán elpusztulnak, ami nem meglepő, ha egy
ilyen fontos feladatot ellátó fehérje hiányzik belőlük. Itt is
megpróbálták mesterségesen kifejeztetni a cyt-c-p vagy a
cyt-c-d géneket és azt tapasztalták, hogy azok a legyek is életképesek
és kikelnek, amelyek a cyt-c-p és azok is, amelyek a
cyt-c-d gént fejezik ki, vagyis mindkét fehérje képes ellátni
az elektronszállítás feladatát is. Gondoljuk csak át: Adott
az ecetmuslicában két citokróm c fehérje, a citokróm c proximal
és a citokróm c distal. Ezek egymástól harminc százalékban különböznek,
körülbelül annyiban, amennyiben az ember és a selyemlepke citokróm
c fehérjéi. Viszont mindkettő képes ellátni az elektronszállítás
feladatát, amely az értelmes tervezés hívei szerint egy "egyszerűsíthetetlenül
összetett" rendszert igényel. Viszont akkor hogy lehet, hogy
ez a rendszer egyszerűsíthetetlenül
összetett, ha az egyik elemének az egyharmada nyugodtam megváltoztatható,
mégis működik? Ezek a rendszerek nem "számos jól illeszkedő,
egymással kölcsönhatásban álló részből állnak"?
Ebben az esetben mégsem illeszkednek olyan jól ezek az elemek? Ez
a példa azért különösen jó, mivel itt minden kísérletet ugyanabban
a szervezetben végeztek, nem beszélhetünk más körülményekről,
más igényekről, más kölcsönhatásokról. Egy "egyszerűsíthetetlenül
összetett" rendszer egyik eleméből az ecetmuslicában két,
egymástól nagymértékben különböző változat található, amelyek
képesek egymást helyettesíteni, egymás működését ellátni. Fontos
kiemelni, hogy ebben az esetben a rendszer többi elemét egyáltalán
nem változtatták meg, egyszerűen egy "alkatrészt" cseréltek
ki egy másikra, mégis működött. Miért van az ecetmuslicának
két különböző citokróm c fehérjéje, ha minden bizonnyal elég
lenne egy is, hiszen mindketten képesek egymás feladatát ellátni?
Emlékezzünk csak: "Egy intelligens cselekvő
számára logikus és egyszersmind hatékony is az
élőlényeket közös biokémiai alapon megtervezni."
Ez az elrendezés egy értelmes cselekvő alkotásának tűnik? Ez
az elrendezés se nem ésszerű, se nem hatékony, ráadásul
teljességgel lehetetlen mögötte bármilyen alkotó
értelem keze nyomát feltételezni.
"Ahogy a képcsövet,
a vezetékeket, az anyacsavarokat és a fémcsavarokat tartalmazza a
tévékészülék, úgy számos fehérje olyan struktúráknak az alkotórésze,
amelyek csak akkor működnek, hogyha valóban minden alkotórész a
helyén van."
"Ha
összehasonlítjuk például egy egérfogóval, amelyben mondjuk
öt alkatrész van és elképzeljük, hogy az egérfogó semmiképpen
sem jöhetett volna létre véletlenek sorozata
által úgy, hogy közben végig működőképes legyen, hanem csak
akkor működik rendeltetésszerűen, amikor már készen van
– hát ki meri feltételezni, hogy a több mint tízezer részből
álló kicsiny sejt-gépezet csak úgy, apró
lépésekkel jött létre, hiszen míg
össze nem állt, nem végezhette el a feladatát?!" (Forrás)
Igaz, hogy még sohasem terveztem
tévét, de feltételezem, hogy ha mondjuk egyharmadával szűkebb,
vagy tágabb anyacsavart használnék hozzá, mint ami éppen oda illik,
nem tudnám összerakni, sőt, ha a képcső egyharmadát változtatnám
meg, akkor se működne a szerkezet. Esetleg ha a tápfeszültséget
nézném el egyharmadával, talán még le is égne, amint áram alá
helyezem. Hogyan lehet hogy egy "egyszerűsíthetetlenül összetett"
rendszer mégis vígan működik tovább, ha az egyik alkotóelemét
egyharmadával megváltoztatjuk? Ha egy egérfogóba egyharmadával
nagyobb vagy kisebb pöcköt szerelünk, ha a lecsapódó keretet, vagy
a rugó erejét mérjük el egyharmadával, ugyanúgy nem működik
majd. Az ecetmuslica citokróm c fehérjéi viszont egyharmadnyi különbségük
dacára is képesek ellátni egymás feladatát. Hát ki meri feltételezni,
hogy ezek jó példák? Az értelmes tervezés hívei által sulykolt
kép, miszerint a mutációs változékonyságnak semmi szerepe sem
lehet biológiai rendszerekben, alapvetően hamis, egyszerű szemfényvesztés.
A biológiai rendszerek általában nagy változékonyságot is elviselnek,
ráadásul egyetlen működést több különböző fehérje is elláthat,
tág tere marad a véletlen mutációs események által létrehozott
változékonyságnak, így a tetszőleges élettelen tárgyakkal dobálózó
hasonlatok egyszerűen nem állják meg a helyüket. Ha már itt tartunk
egy apró kis kérdés: A tévé tojásokat rak? Elevenszülő, netán
osztódik? Vagy az egérfogó? Esetleg a szintén gyakori példa, a
laptop? Ezen élettelen tárgyak nem képesek önmaguk sokszorosítására,
amire az élőlények viszont igen, így teljességgel félrevezető
példaként állításuk. Természetesen senki sem gondolja, hogy
egy laptop magától nőtt volna egy pocsolyába,
ám egyszerűen azért, mert nem rendelkezik azzal az egyszerű
képességgel, hogy önmagáról kisebb-nagyobb hibákkal másolatot
készítsen, nem azért, mert olyan bonyolult eszköz, amely
összetettségi fok már nem jöhet létre véletlen folyamatok
által. Ugyanígy, ha egy almát találunk egy pocsolyában, valamiért
nem feltételezzük, hogy azt éppen most teremtette oda az értelmes
tervező. Még csak azt sem, hogy valaki odatette. Miért? Az is van
olyan bonyolult szerkezet, mint a laptop sőt, sokkal bonyolultabb,
viszont egyetlen almafa nagy mennyiségben képes almát teremni, ezek
közül egy nyugodtan leeshetett, kigurulhatott a fa alól, be a pocsolyába,
kizárólag véletlen folyamatok hatására. Nem lehet, hogy mégis
van némi különbség?
Úgy látszik, ismét nem sikerült
meggyőzni az értelmes tervező létéről. A boncolgatott cikk ismét
csak az evolúció elméletének félremagyarázásán alapul, "felfedezései"
egyszerűen a biológia eredményeinek nem-ismeretén és félreértelmezésén
alapulnak. A cikk írójának feltételezése alapján az evolúció
elmélete azt jelenti, hogy az éppen a kertben ugráló béka a néhány
millió éve kihalt dinoszauruszok őse, illetve az éppen most a laborban
repkedő selyemlepke az ember őse. Ez sajnos időutazás nélkül lehetetlen,
ezt ép eszű ember nem feltételezheti, értelmes tervezővel avagy
nélküle. Azonban ennek a sohasem létezett elméletnek a megcáfolása
sajnos nem érv az evolúció ellen.
Arama E, Bader M, Srivastava
M, Bergmann A, Steller H. (2006): The two Drosophila cytochrome C proteins
can function in both respiration and caspase activation; EMBO J. 25(1):232-43



 "Egy szigorúan szárazföldi állat, amely kezdetben táplálékáért vadászva csak alkalomadtán merészkedett sekély vízbe, majd patakokba és tavakba, végül egy teljesen vízi élőlénnyé alakult, amely nem riadt vissza a nyílt óceántól sem."
"Egy szigorúan szárazföldi állat, amely kezdetben táplálékáért vadászva csak alkalomadtán merészkedett sekély vízbe, majd patakokba és tavakba, végül egy teljesen vízi élőlénnyé alakult, amely nem riadt vissza a nyílt óceántól sem."
 A Puijila fellelési helye azért is fontos, mert pontot tehet az úszólánúak kialakulásának helyét firtató viták végére. Korábban már felmerült, hogy valahol errefele élhetett ennek a csoportnak az őse, de mindmáig, fosszíliák hiányában ezt sokan nem vették komolyan.
A Puijila fellelési helye azért is fontos, mert pontot tehet az úszólánúak kialakulásának helyét firtató viták végére. Korábban már felmerült, hogy valahol errefele élhetett ennek a csoportnak az őse, de mindmáig, fosszíliák hiányában ezt sokan nem vették komolyan.
 A "
A " A "
A " A fejlődés korai szakaszában, mint minden gerinces embrióban, bennünk is könnyen felismerhetőek
A fejlődés korai szakaszában, mint minden gerinces embrióban, bennünk is könnyen felismerhetőek 

Utolsó kommentek