 Bár a növények (részben hozzánemértésünk miatt) szembeötlően kevés figyelmet kaptak eddig a blogban, most részleges kivételt tennék, pont egy egyébként eléggé indulat-generáló, bioetikai kérdés kapcsán.
Bár a növények (részben hozzánemértésünk miatt) szembeötlően kevés figyelmet kaptak eddig a blogban, most részleges kivételt tennék, pont egy egyébként eléggé indulat-generáló, bioetikai kérdés kapcsán.
Az Indexen hétfőn reggel jelent meg egy interjú Horváth Andrással, az MTA Ökológiai és Botanikai Kutatóintézetének munkatársával, amelyben a génmódosított növények (GMO-k) elterjedésének veszélyeiről beszélgetnek Bodoky Tamással. S bár számos érdekes gondolat is elhangzik, végső soron sajnos sem az interjúalanynak, sem a kérdezőnek nem sikerült továbbjutnia a legalapfokúbb GMO-bashingen.
Nota bene, nem azt akarom ezzel mondani, hogy a GMO-k kvázi kritikátlan szeretete lenne az egyetlen üdvözítő és járható út az élet mezején, de nehezen látom annak a logikáját, hogy az objektivitás erőteljes mellőzése miképpen válik a GMO-szkeptikusuk hasznára. Létezik ugyanis egy értelmesen vállalható középút a GMO-k kultikus imádata és a tőlük való, néha már-már irracionálisnak tűnő félelem között, s ahhoz, hogy ezt belássuk, még csak nem is kell a technológia apostolának (mint pl. Dudits Dénes) lennünk. Persze azt azért nem árt tisztázni, hogy mit is csinál(ná)nak a GMO-kat előállítók és milyen cél(ok)ból.
Alapvetően a GMO-fejlesztés (ha egy egész rövid ideig ragaszkodunk az elvi szinthez) nem szól másról, mint amit az emberiség durván 12,000 éve egyébként is lelkesen csinál: többet, nagyobbat és ha ezek mellett még esélyes (és szükséges), jobb ízűt igyekszik egyes növényekből termeszteni. A mennyiségi (és esetenként minőségi) javulás utáni eltökéltséget nem kell alábecsülni, hiszen nemcsak az ún. haszonnövényekkel ajándékozott meg bennünket (lásd pl. búza, kukorica, rizs), de jelentősen hozzájárult egyes növények globális elterjesztéséhez is - a paradicsomtól a kávéig, a gyapottól a padlizsánig, számos növény ma ezer kilométerekre termesztődik eredeti hazájától, és hát a kimcsi sem lenne az, ami, ha a paprika Amerikából, portugál kereskedőkön át nem jut el Koreába. De amíg a korai növénynemesítők technikai repertoárja meglehetősen szűkös volt és kimerült a láthatóan rokon fajok közötti keresztezési kísérletekben, illetve a hasznos mutánsok "vadászatában", a 20. század, konkrétabban a DNS felfedezése, illetve a manipulálását lehetővé tevő eljárások feltűnése, egy radikálisan új megközelítést tett lehetővé: a növényi genom közvetlen megváltoztatását.
Ránézésre az új megközelítés előnye megkérdőjelezhetetlen. Szárazságtűrő fajt akarok? Pikk, egy sivatagi növény "szárazságtűrő" génjét átteszem az általam favorizált fajba. Zavarnak a magok? Pakk, kiütöm a magképzéshez nélkülözhetetlen fejlődési útvonalat. Elegáns és egyszerű. Pontosabban túlontúl egyszerű, hiszen a dolog marhára nem így működik. Mégpedig azért nem, mert ha már nem is egy fekete dobozként tekintünk a növényi sejtre, azért ismereteink messze vannak a tökéletestől. És épp ezért a kívánt gén beültetése után valójában éppen úgy csak drukkolhatunk, hogy jól süljenek el a dolgok, mint a száz évvel ezelőtti növénytermesztők - és ugyanúgy, mint ahogy anno a káposzta és a retek keresztezéséből káposzta levelű és retek gyökerű csodazöldséget várók koppantak, ma sem árt felkészülni arra, hogy a génünk ilyen-olyan okból nem fogja azt tenni, amit elvártunk tőle. Vagy, esetleg nemcsak azt teszi amit szeretnénk, hanem váratlan látható vagy láthatatlan mellékhatásai is vannak. Utóbbiak nem jelentenek nagy gondot, ha pl. egy csak csodálásra való kék rózsáról van szó, viszont tápnövények esetén akár a kísérlet (szomorú) végét jelezhetik: kevés cikibb van, mint ha a megnőtt proteinmennyiség mellett a növény valami mérgező anyagot is rejt. Persze ezt, ugyanúgy, mint a gyógyszerek esetebén, megfelelő kontrollokkal lehetne és kellene vizsgálni, és csak azt engedni forgalomba, amit a soklépcsős procedúra emberi fogyasztásra alkalmasnak talál. Illetve néha azt sem, mert az is előfordulhat, hogy a gén pont azt, és pont úgy csinálja, mint ahogy a tervezőasztalon a növénybiotech-szakik kitalálták, de a külvilágba kikerülve, amolyan "pillangó effektus" révén mégis egész ökoszisztémákat dönthet romba (hiszen a pokolba vezető út is jó szándékkal, mint tudjuk).
Az utóbbi kategóriába tartozhat (azért a feltételes mód, mert a pontos hatás még bőszen vitatott), a legtöbb GMO-ról szóló párbeszéd alfája (és gyakran omegája), az ún. Bt-kukorica. A "Bt" ez esetben nem valami gazdasági társulásra utal, hanem egy baktérium, a Bacillus thuringiensis nevének a röviditése, ui. a transzgénikus kukoricák ez esetben az említett prokarióta egyik toxinját fejezik ki. Ez a kukoricát gyakran károsító rovarok lárváiban kristályosodik ki, végső soron elpusztítva őket. A nagy kérdés persze, hogy a környéken élő többi rovarfajra milyen hatással van az ilyen transzgénikus kukorica, illetve reális-e annak a veszélye, hogy a "beépített" gén valami módon átkerül a környéken élő egyéb növényfajokba (ennek valóban van elméleti lehetősége, az ún. horizontális géntranszfer révén, de hogy konkrétan a gyakorlatban megvalósulhat-e, az már fogasabb kérdés), katasztrófa szélére sodorva az összes környéken élő növényevő rovart, s így közvetve a rájuk épülő táplálékpiramisokat is. A válasz egyelőre bizonytalan, no nem azért, mert nem tanulmányozzák a kérdést, hanem mert egy kicsit vegyesek a tapasztalatok. Tavaly nyáron a Science-ben jelent meg egy metaanalízis, amelyben az eddigi eredményeket és tapasztalatokat próbálták összegezni: a Bt-kukorica (vagy -gyapot) ültetvényeken több rovar található, mint a kontrollként működő, rovarirtóval kezelt "hagyományos" kukoricaföldeken, viszont van néhány rovarcsoport, amelyek képviselői átlagosan kevesebben vannak.
A rovardiverzitás feletti aggódás laikusok számára bizarrnak tűnhet, de nem az, mert a diverzitás csökkenése azt is jelenti, hogy az adott ökológiai rendszer kevésbé tud alkalmazkodni majd a hirtelen változásokhoz. Épp ezért indokoltak a hatástanulmányok, hogy ezek alapján mérlegelhessük, lenne-e annyi haszna adott GMO-faj bevezetésének, hogy az ellensúlyozza a károkat. Megint másik kérdés, hogy a Bt-kukorica és a másik "mumus" a gyomirtószer rezisztens takarmánynövények nem képviselnek mást, csak egy újabb stációt a kártevők elleni harcban, és mivel az "evolúciós fegyverkezési versenyek" állandóak, szinte biztosak lehetünk abban, hogy akárcsak az egykor csodaszernek kikiáltott DDT, max. néhány évnyi előnyt biztosítanak, majd megjelennek (helyenként már megjelentek) a Bt-rezisztens rovarok és a gyomirtó-rezisztens növényi kártevők. Szerencsére ma már jobban tudatában vagyunk az effajta problémának mint a DDT korában, így sok helyen a transzgénikus növényeket kötelező keverni a nem-transzgénikusokkal, így biztosítva, hogy a rezisztens rovarok ne tudjanak gyorsan elterjedni a környéken (a rezisztencia kialakulása gyakran valamilyen más, kisebb élettani hátránnyal jár, így a nem rezisztenciát génjeikben nem hordozó ízeltlábúak szaporodási előnyt élveznek a hordozókkal szemben, ezzel ellensúlyozva azt, hogy ők csak bizonyos (a nem transzgénikus) növényeken tudnak táplálkozni). A módszer sikere egyelőre kétséges, ezért számos szkeptikus (köztük e sorok írója) úgy gondolja, hogy célravezetőbb lenne a "fegyverkezési verseny" biológiai logikáját figyelembe véve keresni a megoldást. Például, ha nem lennének hatalmas kiterjedésű, monokultúrás termőföldek, egy-egy specialista kártevő csak kisebb számban tudna elterjedni, márpedig kevesebb kártevőben kisebb valószínűséggel jelenhet meg a rezisztenciát jelentő mutáció.
A fenti felsorolás azonban csak az érem egyik oldala (egy bővebb terjedelmű kifejtésben persze idetartozna az is, hogy a GMO-knak az sem használt, hogy az őket nyomató vállalatok, pl. a híres-hírhedt Monsanto, gyakran megkérdőjelezhető módon igyekezett maximálni a profitját, de ez egy másik etikai kérdés). Hiszen a gének bevitele nem csak kártevő rezisztens fajokat hozhat létre, hanem olyanokat is, amelyek korábbi tápanyagaik mellett egyéb, az egészséges táplálkozáshoz nélkülözhetetlen anyagokat termelnek. A leghíresebb példa erre az "aranyrizs", amely az A-vitamin előállításához szükséges béta-karotint termel. Ez a növény elvileg évi 1-2 millió ember életét menthetné meg, ugyanis ennyien halnak meg világszerte (elsősorban Afrikában és DK-Ázsiában) A-vitamin hiány miatt. Gyakorlatilag azonban nem több tudományos kuriózumnál, hiszen a Greenpeace és más GMO-ellenes csoportok sikeresen meggátolták a bevezetését. Az ellenkezés elsődleges oka a technológiával szembeni totális bizalmatlanság, illetve az attól való félelem, hogy az aranyrizs ékként tör utat a többi, talán kevésbé létfontosságú GMO bevezetése előtt.
Pedig az aranyrizs és a Dudits által is említett szárazságtűrő fajok kifejlesztése emberek millióinak a megélhetését tehetné biztosítottá, a múlt század közepén lezajlott "zöld forradalom" aktualizált változatát jelenthetné. Nem véletlen, hogy az éhezők megsegítéséért kampányolók közül sokan egyáltalán nem szimpatizálnak a Greenpeace álláspontjával. Vélhetőleg a jelenlegi patthelyzet felett érzett frusztráció késztette a brit libdemek egyik képviselőjét, Dick Taverne-t, hogy igen indulatos (és csöppet sem elfogulatlan) cikket írjon a Prospect magazin novemberi számába. A vádaskodó irományban azonban számos fontos érv szerepel és az egyik kapcsán visszakanyarodnék a Horváth interjú legvégéhez, ahol az interjúalany az organikus gazdálkodást reklámozza.
Az organikus zöldség-gyümölcs finom, sőt gyakran ízesebb mint az "üzemibb" verzióban termesztett változata. De amikor szinte teljesen a természet kezeire hagyatkozunk, akkor pont az előbb említett "zöld forradalom" azon vívmányait vetjük el magunktól, amelyek fél évszázada a megnövekedett hozamot lehetővé tették. Az organikus termelésnek természetesen megvan a maga helye és lehetősége, de ne feledjük: ha a föld teljes lakosságát így akarnánk ellátni, háromszor ennyi termőföldre lenne szükségünk...
A fenti sorokban, amennyire tudtam, megpróbáltam összefoglalni, hogy mi az amire figyelnünk kell a GMO-k termesztése kapcsán és mi az amit elérhetünk velük. Megfelelő ellenőrzéssel ez is egyike lehet az emberiség nagy lehetőségeinek a 21. században. Az szigorú élettani és ökológiai kontroll szükséges és elégséges feltétele kellene legyen az emberiség és GMO-k együttélésnek. Dacból vagy divatból tiltakozni az egész technológia ellen pedig badarság. Mert olyanná válhatunk, mint Károly herceg, amikor a nanotechnológiától a "szürke-goo" okán akarta védeni az emberiséget: nevetségessé.

 A második világháború után, a japán frontról visszatérő Ed Lewis újult erővel vetette bele magát a Drosophila genetikában, a Caltechen levő laborjában. Miközben a homeotikus mutációkat vizsgáló, később Nobel díjjal honorált projektjén dolgozott egy érdekes problémával találta magát szemközt: ha két, látszólag egyforma (vagy nagyon-nagyon hasonló) mutánsom van, akkor miképp állapíthatom meg, hogy a mutációk azonos génben vannak, vagy csak ugyanazon folyamat két, különböző génjében. A probléma megoldása végett fejlesztette ki Lewis az ún. genetikai komplementációt, amelyet logikai egyszerűsége és eleganciája azóta is a genetikusok kedvelt kísérletévé tesz.
A második világháború után, a japán frontról visszatérő Ed Lewis újult erővel vetette bele magát a Drosophila genetikában, a Caltechen levő laborjában. Miközben a homeotikus mutációkat vizsgáló, később Nobel díjjal honorált projektjén dolgozott egy érdekes problémával találta magát szemközt: ha két, látszólag egyforma (vagy nagyon-nagyon hasonló) mutánsom van, akkor miképp állapíthatom meg, hogy a mutációk azonos génben vannak, vagy csak ugyanazon folyamat két, különböző génjében. A probléma megoldása végett fejlesztette ki Lewis az ún. genetikai komplementációt, amelyet logikai egyszerűsége és eleganciája azóta is a genetikusok kedvelt kísérletévé tesz.
 Az elvi részen túl persze még azt sem tudjuk pontosan, hogy az egyes halakban milyen mutáció okozza a szem elsorvadását, mindenesetre mivel számos vak Astyanax populáció nagyon távol él egymsától, feltételezhető, hogy a párhuzamos konvergens evolúció klasszikus példájával állunk szemben, vagyis a vakság egymástól függetlenül alakult ki. Ennek tesztelésére tökéletesen alkalmas a komplementáció, épp ezért végezte ezt el az NYU egyik kutatója, Richard Borowsky.
Az elvi részen túl persze még azt sem tudjuk pontosan, hogy az egyes halakban milyen mutáció okozza a szem elsorvadását, mindenesetre mivel számos vak Astyanax populáció nagyon távol él egymsától, feltételezhető, hogy a párhuzamos konvergens evolúció klasszikus példájával állunk szemben, vagyis a vakság egymástól függetlenül alakult ki. Ennek tesztelésére tökéletesen alkalmas a komplementáció, épp ezért végezte ezt el az NYU egyik kutatója, Richard Borowsky.
 A napokban új, ingyenesen elérhető havilap indult az evolúció iránt érdeklődő laikusok, és a tárgyat oktató tanárok számára,
A napokban új, ingyenesen elérhető havilap indult az evolúció iránt érdeklődő laikusok, és a tárgyat oktató tanárok számára,  Indiánokról írni általában véve hálás feladat, hiszen a téma természeténél fogva, mindig akad egy kis egzotikum, vadregényes tájak, számunkra furcsa kinézetű és viselkedésű emberek. És mint nagy írónk ;-)) May Károly esete is bizonyítja, még csak el sem kell utazni az amerikai kontinensre, hogy a témáról bestsellereket írjunk.
Indiánokról írni általában véve hálás feladat, hiszen a téma természeténél fogva, mindig akad egy kis egzotikum, vadregényes tájak, számunkra furcsa kinézetű és viselkedésű emberek. És mint nagy írónk ;-)) May Károly esete is bizonyítja, még csak el sem kell utazni az amerikai kontinensre, hogy a témáról bestsellereket írjunk.

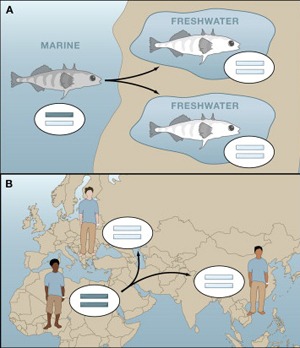 Az elmúlt év legvisszatérőbb témája az állati szőrszínezet, vagyis pigmentáció evolúciója volt, így talán stílszerű, hogy az év utolsó postjának tervezett, de persze az új évbe átcsúszó beírás is ezzel a témával foglalkozzon.
Az elmúlt év legvisszatérőbb témája az állati szőrszínezet, vagyis pigmentáció evolúciója volt, így talán stílszerű, hogy az év utolsó postjának tervezett, de persze az új évbe átcsúszó beírás is ezzel a témával foglalkozzon.
Utolsó kommentek