
Az ehavi Nature Reviews Genetics melléklete az állatok tetszerveződésével kapcsolatos alapkérdéseket taglalja
(leginkább evolúciós szempontból). Ezek tipikusan olyan kérdések, amely
nagyon sok biológus oldalát furdalják, de nehéz kutatásukra pénzt
szerezni, mert szinte lehetetlen komolyan megindokolni, hogy a puszta
ismereten kívül milyen gyakorlati haszna lehet az így megszerzett
tudásnak. Az sem fokozza az ilyenirányú kutatások támogatását, hogy
gyakran obskurus, primitív állatok vizsgálatáról van szó, amelyek nem
igen jönnek szóba mint bármilyen emberi betegség modellállata.
Na jó, akkor ennyi rinyálás után lássuk a medvét. A cikksorozat utolsó darabja Mark Martindale
tollából származik és a többsejtűek tengelyeinek kialakulásával ill. az
ezzel kapcsolatos tulajdonságok megjelenésével foglalkozik ("The
evolution of Metazoan axial properties"). Amiért különösen érdekes, az
nem is az, hogy egy remek összefoglalót ad azokról a tulajdonságokról,
amelyek a legkülönbözőbb állatokban közösek, hanem hogy világosan
megfogalmazza azokat a nagy kérdéseket amelyeket még nem értünk
teljesen vagy egyáltalán.
Jöjjön előbb a már említett közös
tulajdonságok közül a két talán legfontosabb, amelyek a hidráktól a
gerincesekig sokfajta fajban megfigyelhetők, ezért feltehetően a
többsejtűek (metazoák) közös ősében jelen voltak:
- - durva általánosításban a megtermékenyített petesejt animális pólusa felel meg az organizmus leendő elülső (anterior) végének, míg a vegetális pólus a leendő hátsó (poszterior) véggel azonosítható;
- - a beta-catenin nevű fehérje sejtmagban való lokalizációja, számos gén expressziója (pl. brachyury, forkhead, gata) és a gasztruláció (az a folyamat, amelynek során az addig kvázi labda alakú embrióból egy kétrétegű "sapka" lesz, az embrió egyik részének az elsődleges testüregbe való betűrődésével - közben létrejön a másodlagos testüreg és később a csíralemezek) helye között szoros összefüggés van, annak ellenére, hogy az animális-vegetális tengelyen igencsak különbözik a gasztruláció helye újszájúakban és ősszájúakban.
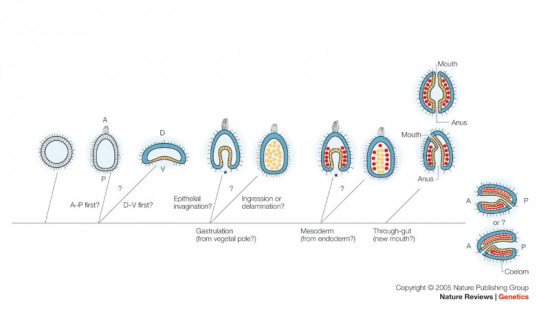
És akkor lássuk a kérdéseket is (pontosabban csak néhányat, mert elég sok van):
- - létezett-e olyan élőlény amelyik az egyszerű blasztula állapotot "testesítette" meg? (Míg a ma ismert legegyszerűbb többsejtű, a Placozoa csoportba tartozó Trichoplax adherens egy többsejtrétegű lapos lemez, amely feltehetően a blastula előtti állapotot tükrözi, az egyszerűségi sorrend következő helyezettjének, a szivacsoknak a lárvái ugyan blastula alkatúak, de a felnőtt egyedek ezekből átalakulással jönnek létre. A kettő közötti átmenetre vonatkozik a kérdés.)
- - az anterior-poszterior (AP) tengely vagy a dorso-ventrális (DV) tengely alakult ki előbb? (Ezek minden kétoldali szimmetriájú állatban fellelhetőek; a dorsális oldal a "hátnak" felel meg, míg a ventrális "hasnak" mind gerincesekben, mind rovarokban, bár egyesek szerint dorsálisként azt az oldalt kellene nevezni, amelyikhez közelebb a központi idegrendszer található - ez pedig a hasi oldal lenne a rovarokban és a háti a gerincesekben.)
- - mikor alakult ki a kétnyílású bélrendszer?
- - mennyire volt a legősibb többsejtű genomja komplex (erre már utaltam a Platynereis kapcsán)? A testalkatok komplexitásának növekedése a gének számának növekedésével, vagy "csak" szabályozásuk és kölcsönhatásaik összetettebbé válásával magyarázható?
- - hogyan és mikor jelentek meg az egyes csíralemezek és az új sejttípusok?
- - lehet-e köze az egysejtű eukarióták polaritásának a többsejtű állatok polaritásához?
- - melyik az ősibb csoport: az ősszájúak vagy az újszájúak? (Az
elnevezés mesterséges és egy korabeli felfogást tükröz, valójában nem
tudjuk, melyik testtípus jelent meg előbb.)
- - létezett-e valaha tökéletesen körkörös szimmetriájú többsejtű állat? (A klasszikus példának tartott csalánozók esetében rendre kiderült, hogy nem minden gén expressziója mutat körkörös szimmetriát, vagyis létezik egy rejtett kétoldaliság.)
- - melyek a legrégebbi gén-kölcsönhatási útvonalak?
Martindale, MQ (2005) The evolution of metazoan axial properties. Nat Rev Gen 6, 917-927.

Utolsó kommentek