
 A tüskés pikó kutatás igazi reneszánszát éli, bár ezúttal a fősodorban nem a halak, egykor Niko Tinbergen érdeklődését is felkeltő viselkedése áll, hanem sokkal inkább a természetben fellelhető formagazdagságuk.
A tüskés pikó kutatás igazi reneszánszát éli, bár ezúttal a fősodorban nem a halak, egykor Niko Tinbergen érdeklődését is felkeltő viselkedése áll, hanem sokkal inkább a természetben fellelhető formagazdagságuk.
A háromtüskés pikó (Gasterosteus aculeatus) legnagyobb természetes populációi a tengerekben élnek, de a kis hal szívósságát mi sem bizonyítja jobban, mint hogy sikerrel megtelepedtek a legutóbbi jégkorszak végén visszavonuló gleccserek helyen kialakuló édesvízi tavakban is. (Az édesvíz és tengervíz ozmotikus tulajdonságai alapvetően különbözőek, ezért nem triviális egy ilyen váltás.) Ráadásul nem is egyszer, hanem nagyon sokszor: a Csendes- és Atlanti-óceán partjai egyaránt tele vannak egyedi tüskés pikó populációknak otthont adó tavakkal (sőt mára már a halnak sikerült elvergődnie Közép-Európába is).
Az alkalmazkodás során a halak külseje is változott: gyakran megfigyelhető a tengeri populációk jellegzetes jegyeinek, a háton levő tüskéknek, a hasúszónak, illetve a testet borító csontpajzsoknak az elsatnyulása (különböző populációkban különböző mértékben). Ennek környezeti okai nem teljesen tisztázottak, de valószínű, hogy része van benne annak, hogy kevesebb kálcium áll rendelkezésre (a csontos kinövések kialakításához, márpedig nélkülözhetetlen ez a vegyület), illetve annak is, hogy a tüskés kinövések, amelyek a tengerben remekül megvédenek a ragadozók ellen (ki lelkesedne egy szúrós kis izé lenyeléséért), a tavakban hátránnyá válhatnak: az ott előforduló ízeltlábú ragadozók számára első rangú fogófelületként működnek.
Ez a természetes formagazdagság elsőosztályú tanulmány-alannyá teszi ezt a halfajt: éppen úgy mint a sok száz drosophilid faj esetében, itt is felvillan annak a lehetősége, hogy az evolúciót működésküzben csípjük fülön, a változatosság genetikai okainak utánajárva. Na persze, annyi hátránnyal szembe kell nézni, hogy a tüskés pikó eddig nem volt a genetikai laborok állandó lakója, s így csak korlátolt ismeretekkel rendelkezünk genomjáról (de napról-napra egyre többel). Viszont ezt a pillanatnyi gyengeséget ellensúlyozza mindaz a tudást, amit más halakról gyűjtöttük össze (elsősorban a zebrahal és a medaka említendő errefele).
 Hogyan lehet tehát annak utánajárni, mi okozza pl. a mellúszók és tüskék elvesztését? Első lépésként keresztezünk egy halat valamelyik tüskés populációból egy tüskétlen közösségből származó társával, majd az így létrejövő, ún. F1 generáció (melynek egyébként minden egyede tüskés, azaz a tulajdonság domináns) egyedeit egymás között szaporítjuk. A keletkező utódok alkotják az F2 generációt, amelyekben már vegyesen lesznek tüskés és nem tüskés egyedek (Mendel törvények, ugye).
Hogyan lehet tehát annak utánajárni, mi okozza pl. a mellúszók és tüskék elvesztését? Első lépésként keresztezünk egy halat valamelyik tüskés populációból egy tüskétlen közösségből származó társával, majd az így létrejövő, ún. F1 generáció (melynek egyébként minden egyede tüskés, azaz a tulajdonság domináns) egyedeit egymás között szaporítjuk. A keletkező utódok alkotják az F2 generációt, amelyekben már vegyesen lesznek tüskés és nem tüskés egyedek (Mendel törvények, ugye).
Második lépésben, olyan molekuláris markereket vetünk be, amelyek viszonylag egyenletesen szétszóródva helyezkednek el a tüskés pikó kromoszómáin, és különböznek egy kicsit a két kiindulási populációban; pontosabban azt vizsgáljuk az F2 generáció egyedeiben, hogy melyik markerek öröklődnek együtt a (tüskés/tüskétlen) tulajdonsággal. Azok a markerek, amelyek minden egyes tüskés F2 halacskában (ill. a tüskés nagyszülőben) megvannak, elég közel kell feküdjenek a tulajdonságot okozó DNS szakaszhoz (és minnél több F2 halat vizsgálunk, annál közelebb).
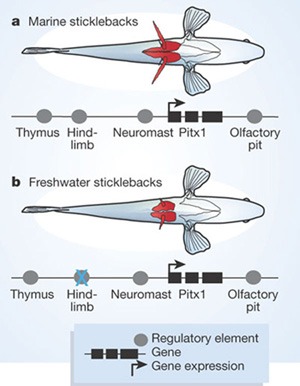
Ezzel az elvvel sikerült leszűkíteni a keresést a pitx1 génre. Ez azért is izgalmas jelölt, mert a hasúszó történetesen a négylábú gerincesek hátsó végtagjának a kvázi homológja, márpedig utóbbi fejlődéséhez elengedhetetlen a Pitx1 jelenléte.
A bökkenő csak az volt, hogy a gén fehérjekódoló része nem tartalmazott egyetlen különbséget sem a tüskés és tüskétlen halak között. Ez azonban csak látszólag probléma, valójában nagyon is várható jelenség: mivel a pitx1-nek sok más szerepe is van a fejlődés során, pl. a szaglószerv, a pajzsmirigy, az oldalvonalszerv kialakításában (más néven egy pleiotróp gén), ha a fehérjekódoló szekvenciáját változtatná meg egy mutáció, akkor az az összes említett szervet érzékenyen érintené. Ha ellenben csak a gén szabályozása módosul, és adott helyen nem expresszálódik, akkor csak az adott helyen zajló fejlődési folyamatokat érinti majd a hiánya.
És éppen ez következett be a pikókban is: a tüskétlen populációkban a pitx1 a leendő hasúszót kivéve, minden más "klasszikus" kifejeződési helyen jelen van, vagyis egyértelmű, hogy egy szabályozó-mutációval állunk szemben (még DNS szinten nem lett beazonosítva, de csak idő kérdése).
Ráadásul több, egymástól távol előforduló - így függetlenül kialakuló - tüskétlen G. aculeatus populációt megviszlantva (sőt, két hasonló kilenctüskés pikó (Pungitius pungitius) populációt is), úgy tűnik, hogy a tüskétlenedéshez a halak igencsak előszeretettel használják fel a pitx1 szabályozó szekvenciáinak változtatgatását. (Hasonló egy kicsit a helyzet, mint a fekete mintázatoknál, ahol az mc1r gén kódoló részének mutációja a ludas sok esetben, csak itt nem a fehérjekódoló részben következnek be a változások.) Sőt, a paleontológiai leletek azt mutatják, hogy a tüskék elhagyása már évtízezredek óta egy kedvelt időtöltése volt a pikóféléknek, lásd ezt az kis videot.
 Akkor már csak a csontlemezek, ill. azok elhagyásának a kérdése maradt. Itt is a fentebb ismertetett elvhez hasonlóan jártak el David Kingsley-ék (akinek a laborjában az ilyen irányú kutatások zöme zajlik manapság) és a fent említett F2 nemzedéknek köszönhetően találtak rá az ectodysplasin (eda) génre.
Akkor már csak a csontlemezek, ill. azok elhagyásának a kérdése maradt. Itt is a fentebb ismertetett elvhez hasonlóan jártak el David Kingsley-ék (akinek a laborjában az ilyen irányú kutatások zöme zajlik manapság) és a fent említett F2 nemzedéknek köszönhetően találtak rá az ectodysplasin (eda) génre.
És ez megintcsak nem egy hétköznapi jelölt, ui. a gén humán mutációi számos emberi ektodermális származék (haj, fogak, izzadtság mirigyek) fejlődési rendellenességével járnak. A pikók csontlemezei szintén ektodermális eredetűek, vagyis a gén funkciója jól megőrződött az évmilliók során. (Egyébként szintén úgy tűnik, hogy egy szabályozó mutációról van szó.) És, hogy a történet kerek legyen: az elmúlt években laboratóriumi mutagenezis kísérletekben olyan medaka és zebrahal példányokat sikerült izolálni, amelyek az EDA aktivitésvesztése miatt pikkelytelenek lettek. A külvilág is egy labor, csak a természet nagyobb egyedszámmal, és lényegesen nagyobb időskálán dolgozik, mint mi, mezei kutatók :-).
Shapiro, MD, Marks, ME, Peichel, CL, Blackman, BK, Nereng, KS, Jónsson, B, Schulter, D, Kingsley, DM (2004) Genetic and developmental basis of evolutionary pelvic reduction in threespine sticklebacks. Nature 428: 717-723.
Colosimo, PF, Hosemann, KE, Balabhadra, S, Villareal, G Jr, Dickson, M, et al. (2005) Widespread parallel evolution in sticklebacks by repeated fixation of Ectodysplasin alleles. Science 307: 1928-1933.
Colosimo, PF, Peichel, CL, Nereng, K, Blackman, BK, Shapiro, MD, et al. (2004) The Genetic Architecture of Parallel Armor Plate Reduction in Threespine Sticklebacks. PLoS Biol 2(5): 635-641. doi:10.1371/journal.pbio.0020109
Shapiro, MD, Bell, MA, Kingsley, DM (2006) Parallel genetic origins of pelvic reduction in vertebrates. PNAS 103: 13753-13758. doi:10.1073/pnas.0604706103

Utolsó kommentek