
 Ha szemügyre veszünk egy ezerlábút, talán az első dolog ami a szemünkbe ötlik, hogy teste ismétlődő egységekből (ún. szegmensekből) áll, amelyek mint megannyi egyforma „építőmodul” sorakoznak egymás mögött. Figyelmesebben szétnézve valami hasonlóra lelhetünk rákokban, rovarokban és más ízeltlábúakban is, sőt, mi magunk, gerincesek sem vagyunk kivételek az efajta modularitás alól, bár esetünkben az ismétlődő egységeket leginkább csak az embrionális fejlődés során lehet könnyen elkülöníteni. S ugyan fontos hangsúlyozni, hogy utóbbi állatcsoportokban, az ezerlábúak szegmenseitől eltérően, az „építőmodulok” nem tökéletesen egyformák, mégis egyértelműnek tűnik, hogy amit látunk az egyfajta „variációk egy témára”, vagyis minden egység leszármaztatható egy szegmens-prototípusból. Ebből persze rögtön adódik a kérdés: milyen mechanizmusok felelősek az egyes szegmens-variánsok kialakulásáért?
Ha szemügyre veszünk egy ezerlábút, talán az első dolog ami a szemünkbe ötlik, hogy teste ismétlődő egységekből (ún. szegmensekből) áll, amelyek mint megannyi egyforma „építőmodul” sorakoznak egymás mögött. Figyelmesebben szétnézve valami hasonlóra lelhetünk rákokban, rovarokban és más ízeltlábúakban is, sőt, mi magunk, gerincesek sem vagyunk kivételek az efajta modularitás alól, bár esetünkben az ismétlődő egységeket leginkább csak az embrionális fejlődés során lehet könnyen elkülöníteni. S ugyan fontos hangsúlyozni, hogy utóbbi állatcsoportokban, az ezerlábúak szegmenseitől eltérően, az „építőmodulok” nem tökéletesen egyformák, mégis egyértelműnek tűnik, hogy amit látunk az egyfajta „variációk egy témára”, vagyis minden egység leszármaztatható egy szegmens-prototípusból. Ebből persze rögtön adódik a kérdés: milyen mechanizmusok felelősek az egyes szegmens-variánsok kialakulásáért? Csak a változatosság örök, mondhatjuk őszintén, hiszen a variáció a bennünket körbevevő élő anyag egyik alapvető jellemzője. Mindenütt és mindig jelen van, nincs két egyed amelyik tökéletesen egyforma lenne. Ebből a változatosságból válogat a természetes szelekció is, azaz pont ezek a kisebb-nagyobb különbözőségek adják az evolúció hajtóerejét. Amint azt a brit William Bateson variációkról szóló, 1894-ben kiadott klasszikus művében megfogalmazta:
“[...] éppen ezért a Variáció, bármi is a kiváltó oka és hatása bármennyire is korlátozott legyen, az Evolúció meghatározó jelensége. A Variáció valójában maga az Evolúció. Éppen ezért a legkézenfekvőbb megoldás az Evolúció problémájának megoldására a Variáció tényeinek a tanulmányozása.”
Bateson bizonyos szempontból a genetika egyik atyjának tekinthető (ő használta először ezt a kifejezést az öröklődéstanra), s a fenti sorok papírra vetésekor még igencsak gyerekcipőben járt az említett tudomány. Mendel törvényei türelmesen vártak újrafelfedezésükre, az örökítőanyag mibenlétére pedig csak több mint fél évszázaddal később került sor és még több időre volt szükség, hogy a genetika eszköztára annyira kibővüljön, hogy az egyes géneket szekvencia szinten tanulmányozni tudjuk. Mindez idő alatt Bateson felismerése semmit sem vesztett aktualitásából, biológus generációk egész sora szentelte idejét és karrierjét annak, hogy feltárja az öröklődés és evolúció szabályait a variációk tanulmányozása révén. Ezeknek a vizsgálatoknak az egyik elterjedt tárgya az egyszerű ecetmuslica, a Drosophila melanogaster lett.
A variációk egyik típusa különösen felkeltette Bateson érdeklődését: a szóban forgó állatok állatok egy-egy adott szegmense valamelyik másik szegmens jellegét vette fel (pl. csápok helyett lábai nőttek, vagy szárnyai ott ahol nem kellett volna). Ezeket homeotikus mutációknak nevezte el, a szegmensek átalakulását pedig homeózisnak. "Ebben a munkában ez a fajta variáció különösen fontos lesz és hiszek benne, hogy a jövőben a jelentősége és előfordulásának módja nagy érdeklődésre tarthat számot”, írta fent említett művében és talán még önmaga is meglepődött volna, ha tudja később mennyire igaza lesz.
Az első homeotikus mutációkat laboratóriumi körülmények között a múlt század elején izolálták, a genetika egy másik kiemelkedő alakja, Thomas Morgan laboratóriumában. Ekkor derült fény arra is, hogy tovább örökíthető, a kromoszómákon levő génekhez kapcsolt tulajdonságról van szó. Arra azonban további fél évszázadig kellett várni, hogy valaki tüzetesebb vizsgálatnak vesse alá a homeotikus mutációkat. Erre végül Morgan munkásságának egyik szellemi örököse, Ed Lewis vállalkozott. Lewis homeotikus mutációk egész seregét izolálta, majd meghatározva pontos helyzetüket a kromoszómákon, megfogalmazta a fejlődésbiológia egyik leghíresebb és egyben legenigmatikusabb szabályát, a kollinearitásét. Ez azt fejezi ki, hogy a homeotikus gének egymás mellett helyezkednek el a kromoszómákon, és kromoszómális helyzetük összefüggésben van azzal, hogy hol, a test mely szegmenseiben fejtik ki a hatásukat: minél előrébb helyezkedik egy ilyen gén el a kromoszómán, annál előrébb található szegmensekre hat és vice versa.
Amikor pár évvel később a géneket is sikerült külön-külön izolálni, arra is fény derült, hogy mindez kapcsolatban van azzal, hogy hol fejeződnek ki a testben, hiszen az előrébb levő géneknek előrébb található az expressziós tartományuk, mint a hátrább helyet foglalóknak. Az is kiderült, hogy a homeotikus gének olyan transzkripciós faktorok, amelyek DNS kötő részét (az ún. homeodomént) egy nagyon hasonló, 180 bázispár hosszú szekvencia jellemzi. Ez utóbbi a homebox, amely után az ilyen géneket gyakran homeobox-géneknek nevezik az irodalomban. Amire azonban szinte senki nem számított, hogy ugyanezen géneket megtalálták egerekben is (sőt azóta a legkülönbözőbb állatcsoportokban), ahol hasonló sorrendben elhelyezkedve, hasonló szerepet töltenek be. A kollinearitási szabály univerzalitása az elmúlt néhány évtized legjelentősebb felfedezéseinek egyike; az, hogy ugyanaz a rendszer felelős minden ma élő, kétoldali szimmetriájú állat hossztengelyének felosztásáért, egyértelműen bizonyítja a közös evolúciós eredetet is.
Már Lewis is felismerte, hogy a homeotikus géneknek kulcsszerepük lehetett az evolúcióban, különös tekintettel az egyes szegmensek közötti különbségek kialakításában. Mint azt egyik, ma már klasszikusnak számító összefoglalásában megfogalmazta:
„A muslicák majdnem biztos, hogy négy és nem két szárnyú rovarokból alakult ki, és maguk a rovarok olyan ízeltlábúakból származnak, amelyeknek több lábuk volt mint hat.” (Megj.: a Drosophila a Kétszárnyúak (Diptera) csoportjába tartozik, ahol a harmadik tor szelvényen levő szárny pár egy, az egyensúlyozásban fontos szervvé, a billérré alakult).
Mai ismereteink alapján már tudjuk, hogy Lewis fején találta a szöget, de mielőtt az általa is említett példákat vennénk szemügyre, tegyünk egy kis kitérőt egy másik ízeltlábú csoport, a rákok felé.
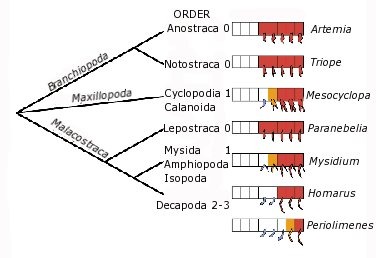 Rákokról beszélve leginkább mindenkinek a folyami rák jut eszébe az ollóival, pedig a rákok osztálya (Crustatcea) az egyik legsokszínűbb ízeltlábú osztály. Más ízeltlábúakhoz hasonlóan az első szelvények végtagjai csápokká illetve szájszervekké módosultak (erről egyszer rövidebben már ejtettem szót), de igen nagy változatosság figyelhető meg abban, hogy pontosan hány végtagnak van szerepe a táplálkozásban. A legegyszerűbb esetekben mindössze három pár: az első az ún. felső állkapcsot (mandibulát), második kettő pedig az alsó állapcsokat (maxillákat) alkotja. Számos esetben azonban ezekhez a következő szelvények lábaiból módosult segédállkapcsok vagy állkapocslábak (maxillipedia - kék függelékek a mellékelt ábrán) csatlakoznak. Ezek száma ami változatos a különböző rákok között. Az apró sóférgek (Artemia) vagy nyári pajzsosrák (Notostraca) esetében például egyáltalán nem lelünk ilyeneket, az ászkarákok (Isopoda) és hasadtlábú rákok (Mysida) esetében egy-egy párat, míg az ismertebb tízlábú rákok (pl. homár - Homarus) esetében 2-3 pár maxillipedia fedezhető fel. Mint arra fény derült, az, hogy mely szelvényeken találhatunk egy pár járólábat, és mely szelvényeken egy pár állkapocslábakat, szorosan összefügg azzal, hogy az adott szelvény embrionális fejlődése során kifejeződik e benne két homeobox-gén, az Ubithorax (Ubx) és az abdominalA (abdA). Ez a két homeotikus gén az ízeltlábúak között általában a hátsó testtáj jellegének kialakításáért felelős (hamarosan még lesz róluk szó) és az említett rákok esetében minél több állkapocslábbal rendelkezik az állat, annál hátrább kezdődik a kifejeződdési tartományuk (piros ill. sárga négyzetek - a sárga szín gzenge expressyiót jelöl). Azaz, azokban a szelvényekben ahol a fejlődés során megjelenik a termékük, ott járólábak alakulnak ki, ahol pedig nem (fehér négyzetek), ott maxillipediumok.
Rákokról beszélve leginkább mindenkinek a folyami rák jut eszébe az ollóival, pedig a rákok osztálya (Crustatcea) az egyik legsokszínűbb ízeltlábú osztály. Más ízeltlábúakhoz hasonlóan az első szelvények végtagjai csápokká illetve szájszervekké módosultak (erről egyszer rövidebben már ejtettem szót), de igen nagy változatosság figyelhető meg abban, hogy pontosan hány végtagnak van szerepe a táplálkozásban. A legegyszerűbb esetekben mindössze három pár: az első az ún. felső állkapcsot (mandibulát), második kettő pedig az alsó állapcsokat (maxillákat) alkotja. Számos esetben azonban ezekhez a következő szelvények lábaiból módosult segédállkapcsok vagy állkapocslábak (maxillipedia - kék függelékek a mellékelt ábrán) csatlakoznak. Ezek száma ami változatos a különböző rákok között. Az apró sóférgek (Artemia) vagy nyári pajzsosrák (Notostraca) esetében például egyáltalán nem lelünk ilyeneket, az ászkarákok (Isopoda) és hasadtlábú rákok (Mysida) esetében egy-egy párat, míg az ismertebb tízlábú rákok (pl. homár - Homarus) esetében 2-3 pár maxillipedia fedezhető fel. Mint arra fény derült, az, hogy mely szelvényeken találhatunk egy pár járólábat, és mely szelvényeken egy pár állkapocslábakat, szorosan összefügg azzal, hogy az adott szelvény embrionális fejlődése során kifejeződik e benne két homeobox-gén, az Ubithorax (Ubx) és az abdominalA (abdA). Ez a két homeotikus gén az ízeltlábúak között általában a hátsó testtáj jellegének kialakításáért felelős (hamarosan még lesz róluk szó) és az említett rákok esetében minél több állkapocslábbal rendelkezik az állat, annál hátrább kezdődik a kifejeződdési tartományuk (piros ill. sárga négyzetek - a sárga szín gzenge expressyiót jelöl). Azaz, azokban a szelvényekben ahol a fejlődés során megjelenik a termékük, ott járólábak alakulnak ki, ahol pedig nem (fehér négyzetek), ott maxillipediumok.
Ez a szabályszerűség azonban nem csak rákokra jellemző. A távoli rokon százlábúak esetében az állkapocslábakat tartalmazó szelvényben szintén nem észlelhető sem az Ubx, sem az abdA kifejeződése.
Az előzőekben láthattuk, hogy a homeotikus gének expressziójának helyszíne miképpen befolyásolhatja az egyes szegmensek identitásának kialakulását. Van azonban arra is példa, hogy a nem a kifejeződés helye változik, hanem a homebox gén funkciója módosul. Erre kínál jó példát Ed Lewis második „megérzése”, miszerint a hat-lábú rovarok több végtagú ízeltlábúaktól származtathatók. S bár a forgatókönyv változik, a főszereplő ugyanaz marad: az Ubx homeobox-gén.
Mint azt néhány sorral feljebb említettem az Ubx és abdA nagy általánosságban megfogalmazva, az ízeltlábúak hátsó testtájának kialakításában játszanak fontos szerepet. A rovarok esetében ez a potroh, amelynek egyik jellegzetessége, hogy szelvényein nincsenek lábak. A rák szájszervek történetének ismeretében az említett gének termékének jelenléte illetve a lábak hiánya között feltűnő a korreláció, de hogy ennek miértjét is megérthessük, egy picit távolabbra kell tekintenünk az ízeltlábúaknál.
 Az őslégcsövesek (Onycophora) első látásra leginkább játékbolti plüsshernyóknak tűnnek, de valójában az ízeltlábúakkal közeli rokonságban álló csoportról van szó, s amennyire megtudjuk ítélni a rendelkezésünkre álló fosszíliák alapján, testfelépítésük alig változott a kambrium óta. Testük ugyanúgy szelvényezett, mint távoli rokonaiké, de a nem borítja őket vastag kitinréteg és minden testszelvényükön jelen levő lábaik sem ízekből épülnek fel. Mindezen tulajdonságaik és távoli rokonságuk miatt azonban remek jelöltek, ha azt szeretnénk vizsgálni, milyenek is lehettek a korai ízeltlábúak homeobox génjei. Az Onycophorák Ubx génje (OUbx) számos hasonló tulajdonsággal bír, mint Drosophila megfelelője (DUbx). Például, ha egy kísérlet során a fejlődő muslica embriók fejébe juttatjuk, akkor mindkettő képes a csápokból lábakat csinálni. De más tulajdonságaikban lényegesen különböznek.
Az őslégcsövesek (Onycophora) első látásra leginkább játékbolti plüsshernyóknak tűnnek, de valójában az ízeltlábúakkal közeli rokonságban álló csoportról van szó, s amennyire megtudjuk ítélni a rendelkezésünkre álló fosszíliák alapján, testfelépítésük alig változott a kambrium óta. Testük ugyanúgy szelvényezett, mint távoli rokonaiké, de a nem borítja őket vastag kitinréteg és minden testszelvényükön jelen levő lábaik sem ízekből épülnek fel. Mindezen tulajdonságaik és távoli rokonságuk miatt azonban remek jelöltek, ha azt szeretnénk vizsgálni, milyenek is lehettek a korai ízeltlábúak homeobox génjei. Az Onycophorák Ubx génje (OUbx) számos hasonló tulajdonsággal bír, mint Drosophila megfelelője (DUbx). Például, ha egy kísérlet során a fejlődő muslica embriók fejébe juttatjuk, akkor mindkettő képes a csápokból lábakat csinálni. De más tulajdonságaikban lényegesen különböznek. 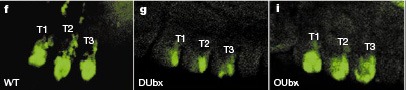 Ha a DUbx gént nagy mennyiségben mesterségesen kifejeztetjük az embrió tor szelvényeiben, azok potroh szelvényekké alakulnak és ennek megfelelően bennük a végtagképződés lelassul. Ellenben az OUbx génnek semmilyen hasonló hatása nincs. Minden azért van, mert az evolúció során, bár maga homeodomén nem változott, a rovarok Ubx génje olyan szekvenciákra tett szert, amelyek a végtagfejlődés egyik kulcsgénjét, a Distal-less-t (Dll) képesek kikapcsolni (lásd mellékelt ábra g. paneljét, ahol a DUbx kifejezése gátolja a torszelvényekben egyébként megvalósuló Dll expressziót). Mivel az őslégcsövesekben ezek a változások nem jelentek meg, az Ubx kifejeződése itt nincs hatással a lábfejlődésre (i). (A teljes tényszerűség okáért, azt meg kell megemlíteni, hogy az OUbx nem jut kifejeződésre az Onycophorák minden szelvényében – még nem tudjuk, hogy miért nem –, csak a leghátsóbbakban – de azokon is vannak lábak.)
Ha a DUbx gént nagy mennyiségben mesterségesen kifejeztetjük az embrió tor szelvényeiben, azok potroh szelvényekké alakulnak és ennek megfelelően bennük a végtagképződés lelassul. Ellenben az OUbx génnek semmilyen hasonló hatása nincs. Minden azért van, mert az evolúció során, bár maga homeodomén nem változott, a rovarok Ubx génje olyan szekvenciákra tett szert, amelyek a végtagfejlődés egyik kulcsgénjét, a Distal-less-t (Dll) képesek kikapcsolni (lásd mellékelt ábra g. paneljét, ahol a DUbx kifejezése gátolja a torszelvényekben egyébként megvalósuló Dll expressziót). Mivel az őslégcsövesekben ezek a változások nem jelentek meg, az Ubx kifejeződése itt nincs hatással a lábfejlődésre (i). (A teljes tényszerűség okáért, azt meg kell megemlíteni, hogy az OUbx nem jut kifejeződésre az Onycophorák minden szelvényében – még nem tudjuk, hogy miért nem –, csak a leghátsóbbakban – de azokon is vannak lábak.)
Folytköv.
Galant, R, Carroll, SB (2002) Evolution of a transcriptional repression domain in an insect Hox protein. Nature 415: 910-913.
Grenier, JK, Carroll, SB (2000) Functional evolution of the Ultrabithorax protein. PNAS 97: 706-709.
Averof, M, Patel, NH (1997) Crustacean appandage evolution associated with changes in Hox gene expression. Nature 388: 682-686.
Lewis, EB (1978) A gene complex controlling segmentation in Drosophila. Nature 276: 565-570.
Duncan, I, Montgomery, G (2002) E.B. Lewis and the Bithorax complex I. Genetics 160: 1265-1272.
Duncan, I, Montgomery, G (2002) E.B. Lewis and the Bithorax complex II. - From cis-trans test to the genetic control of development. Genetics 161: 1-10.

Utolsó kommentek