
 A lábak számának csökkenése még nem minden, hiszen rovarok evolúciója során az egyik legjelentősebb lépés a szárnyak kialakulása és ezzel együtt a repülés megjelenése volt. Utóbbinak fontos szerepe volt abban, hogy a későbbiekben az állatvilág egyik legsikeresebb és legnépesebb állatcsoporttá váltak. Hogy miként alakultak ki a szárnyak, még nem tudjuk pontosan, de a fosszilizálódott ősi rovarok alakja, valamint a környezetünkben előforduló kortársaikból származó genetikai információi jó kiindulási pontot kínál arra, hogy egy hipotézist állítsuk fel erre vonatkozólag.
A lábak számának csökkenése még nem minden, hiszen rovarok evolúciója során az egyik legjelentősebb lépés a szárnyak kialakulása és ezzel együtt a repülés megjelenése volt. Utóbbinak fontos szerepe volt abban, hogy a későbbiekben az állatvilág egyik legsikeresebb és legnépesebb állatcsoporttá váltak. Hogy miként alakultak ki a szárnyak, még nem tudjuk pontosan, de a fosszilizálódott ősi rovarok alakja, valamint a környezetünkben előforduló kortársaikból származó genetikai információi jó kiindulási pontot kínál arra, hogy egy hipotézist állítsuk fel erre vonatkozólag.
A legősibb rovaroknak, amelyek leginkább a ma is élő pikkelykéhez hasonlítottak, egyáltalán nem volt szárnya. Emiatt természetesen csak kétféle élettér közül választhattak: vagy a szárazföldön, vagy a vízben éltek. A repülő rovarok őse feltehetően az utóbbiak közé tartozott, s a rákokhoz hasonlóan az oxigén felvételt kopoltyúkon keresztül valósította meg. A kopoltyúk az állat testének majd minden szegmensén jelen lehettek, tollszerű kitüremkedések formájában, hasonlóan a kérészek lárváihoz. Ezek a kitüremkedések aztán fokozatosan egyre nagyobbak és szilárdabbak lettek, s végül alkalmasakká válhattak, hogy segítségükkel gazdájuk, egy már kevésbé a vízhez kötött életciklusa során vitorlázórepülőhöz hasonlóan irányítsa ugrását. Végül, az aktív repülés kialakulásával párhuzamosan ezek a kitüremkedések eltűntek a hátsó szelvényekről, mert néha a kevesebb több: amíg nagyon sok szárny nem képes jól irányítható repülésre, egy-két pár tökéletesen megfelel a célnak.
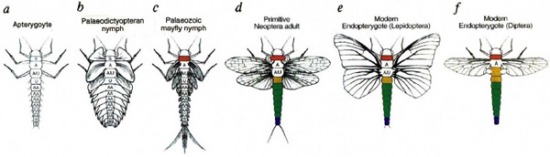
A szárnyak kopoltyú eredetét látszik bizonyítja az ősi rovarok szárnyainak bő erezettsége is, de ami ennél is jobban ezt az evolúciós utat támasztja alá, az az a megfigyelés, hogy a szárnyak fejlődése során ugyanazok a gének (pdm és apterous) játszanak kulcsszerepet, mint a rákok kopoltyújának kialakulásakor.
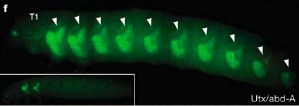 Talán, nem okozok nagy meglepetést, ha elárulom, hogy a szárnyak elvesztése is minden valószínűség szerint egy homebox gén működéséhez kapcsolódik. Nem is akármelyikéhez: mivel a rovaroknak csak két hátsó torszelvényükön van szárny, azaz olyan szelvényeken, ahol lábaik is vannak, a gyanú szinte egyből az Ubx-abdA párosra terelődött. Mint kiderült, egyáltalán nem alaptalanul, hiszen azokban a muslica embriókban amelyekből mindkét gén hiányzik, nemcsak a két hátsó torszelvényben, de az összes potrohszelvényben is észrevehető a szárnyképző program beindítása. És ez igaz más rovarokra is, pl. a lisztbogárra - utóbbinak a lárvája látható jobboldalon: az említett génpáros nélkül a második tor szelvénytől kezdve minden tor- és potrohszelvényen jelen vannak a szárnykezdemények (egy "normális", kezeletlen lárván, mint a bal alsó sarokban csak a második és harmadik torszelvényen láthatunk hasonlót).
Talán, nem okozok nagy meglepetést, ha elárulom, hogy a szárnyak elvesztése is minden valószínűség szerint egy homebox gén működéséhez kapcsolódik. Nem is akármelyikéhez: mivel a rovaroknak csak két hátsó torszelvényükön van szárny, azaz olyan szelvényeken, ahol lábaik is vannak, a gyanú szinte egyből az Ubx-abdA párosra terelődött. Mint kiderült, egyáltalán nem alaptalanul, hiszen azokban a muslica embriókban amelyekből mindkét gén hiányzik, nemcsak a két hátsó torszelvényben, de az összes potrohszelvényben is észrevehető a szárnyképző program beindítása. És ez igaz más rovarokra is, pl. a lisztbogárra - utóbbinak a lárvája látható jobboldalon: az említett génpáros nélkül a második tor szelvénytől kezdve minden tor- és potrohszelvényen jelen vannak a szárnykezdemények (egy "normális", kezeletlen lárván, mint a bal alsó sarokban csak a második és harmadik torszelvényen láthatunk hasonlót).
A muslicák, a házilegyekkel egyetemben a kétszárnyúak (Diptera) rendjébe tartoznak, azaz csak az második torszelvényükön (T2) lelhetünk klasszikus, hártyás szárnyakat, a harmadikon (T3) levő páros különleges egyensúlyszervvé, úgynevezett billérré alakult. Az első homeotikus mutánsok egyike, amelyeket mint korábban említettem Thomas Morgan laborjában izoláltak, azonban pont egy négy szárnyat viselő egyed volt, amelynek billérjei helyén is szárnyak voltak, azaz a harmadik torszelvénye a második szakasztott másolata lett. 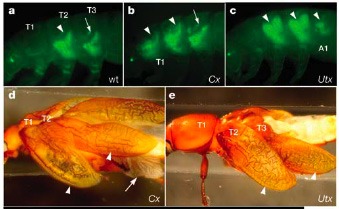 A megkettőződő torszelvény miatt, a mutációt bithorax-nak keresztelték, s ma már azt is tudjuk, hogy az Ubx egyik szabályozó régiója változott meg. Hasonló hatás figyelhető meg a lisztbogarakban is. Ezekben a T2-n található kemény fedő-szárny „duplázódik” meg, ha kiütjük a rovarban az Ubx-t (amit ez esetben Utx-nek kereszteltek), amely normális esetben alacsony szinten jelen van T3-ban.
A megkettőződő torszelvény miatt, a mutációt bithorax-nak keresztelték, s ma már azt is tudjuk, hogy az Ubx egyik szabályozó régiója változott meg. Hasonló hatás figyelhető meg a lisztbogarakban is. Ezekben a T2-n található kemény fedő-szárny „duplázódik” meg, ha kiütjük a rovarban az Ubx-t (amit ez esetben Utx-nek kereszteltek), amely normális esetben alacsony szinten jelen van T3-ban.
Mindeddig csak a hátsó két torszelvényről beszéltem, de mi a helyzet az első torszelvénnyel? Itt egy másik homeobox gén a, a sex-combs reduced (scr vagy Cx), kapcsolja ki a szárnyképződés programját a fejlődés közben. Ha például lisztbogarakban kikapcsoljuk a gén expresszióját, az első torszelvényen is létre jönnek a szárnyak.
Összefoglalva tehát, a valamikori sokvégtagú ízeltlábú-ősből a mai legyekig tartó evolúciós út, ha nem is triviális, de viszonylag egyszerű szabályokkal írható le. A folyamat során leggyakrabban néhány homeobox-gén kifejeződési helye változott meg, vagy (ritkábban) a funkciójuk módosult.
Tomoyasu, Y, Wheeler, SR, Denell, RE (2005) Ultrabithorax is required for membranous wing identity in the beetle Tribolium castaneum. Nature 433: 643-647.
Averof, M, Cohen, SM (1997) Evolutionary origin of insect wings from ancestral gills. Nature 385: 627-630.
Carroll, SB, Weatherbee, SD, Langeland, JA (1995) Homeotic genes and the regulation and evolution of insect wing number. Nature 375: 58-61.

Utolsó kommentek