
 (Szerencsére SexComb nincs vakáción, így egy kis szünet állhat be a poszt-ínséges időkben. - dolphin)
(Szerencsére SexComb nincs vakáción, így egy kis szünet állhat be a poszt-ínséges időkben. - dolphin)
A tervezéselmélet egyik fő érve, amit úton-útfélen előhoznak, az egyszerűsíthetetlen összetettség, ami a hívek szerint egyértelműen bizonyítja az élőlények tervezett voltát.
Maga az érv elég régi, de az ÉRTEM által használt mai formájának alapjait Michael Behe a Darwin fekete doboza című könyvében fektette le, az eredeti meghatározása valahogyan így hangzott:
"A single system which is composed of several interacting parts that contribute to the basic function, and where the removal of any one of the parts causes the system to effectively cease functioning."
"Egységes rendszer, amely számos egymással kölcsönható részegységből áll, amelyek mind hozzájárulnak az alapvető működéshez és ahol a részegységek bármelyikének az eltávolítása a rendszer működésképtelenségét eredményezi."
Az érvelés annyi, hogy ezek a rendszerek nem alakulhattak ki apró változások sorozatával, hiszen minden részegységük szükséges a működésükhöz. "Egyszerűsíthetetlen összetettség nem állhat elő fokozatosan, egy korábbi rendszer jelentéktelen, egymás utáni változásainak eredményeként, mert definíció szerint egy egyszerűsíthetetlenül összetett rendszernek bármely őse működésképtelen."
Behe meg is nevezett pár ilyen rendszert, az eredeti példái közül most a baktériumok ostorával szeretnék részletesebben foglalkozni, mivel magyar nyelven erről írtak eddig a legkevesebbet.
"A baktérium ostora" alatt az Escherichia coli baktérium ostorát értjik általában, amely valóban számos egymással kölcsönható részegységből épül fel, amelyek eltávolítása működésképtelenné teszi az E. coli baktérium ostorát. Ennyiben teljesülnek Behe feltételei, viszont, ha megnézzük, miből is áll a többi baktérium ostora, már merőben más képet kapunk. Szerencsére találtam egy összefoglaló cikket (Pallen M. J. és Matzke N. J. 2006.), így ennek a táblázatát venném át:
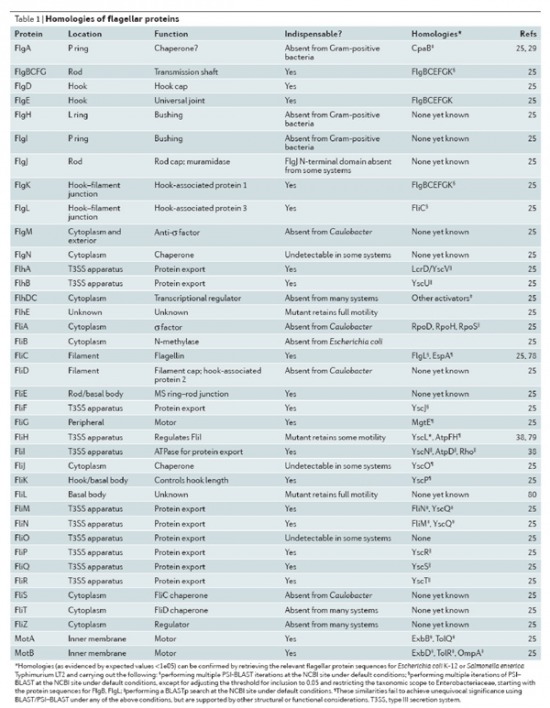
Mit is látunk ezen a táblázaton? Léteznek más baktériumok, amelyek ostora eltérő felépítésű, mint az E. coli-é, ezekből hiányoznak olyan részegységek, amelyek az elsőként vizsgált ostor működéséhez elengedhetetlenek. Sőt, léteznek olyan baktériumok, amelyek ostora az E. coli-énál több alkatrészből áll. Akkor ez hogy van? Mégsem egyszerűsíthetetlenül összetett ez a rendszer? Nézzük meg például az FlgI fehérjét! Ez szükséges az E. coli ostor működéséhez, ha az ezt kódoló gént eltávolítjuk a genomból, az E. coli ostora nem működik (Hizukuri és munkatársai 2006.). Ez eddig egybevág Behe elméletével, ám ha megnézzük, azt találjuk, hogy ez a fehérje hiányzik az összes Gram-pozitív baktériumból. Akkor ez hogy lehet? Van egy egyszerűsíthetetlenül összetett rendszernek egy olyan változata, amelyből hiányzik az egyik alkotórész, mégis működik! Mégiscsak lehetnek működőképes elődei, amelyek nem tartalmazzák minden részegységét, például ott a Gram-pozitív baktériumok ostora! Ugyanígy végigmehetünk a táblázat többi során is és láthatjuk, hogy bizony az "egyszerűsíthetetlenül összetett" rendszer több alkotóeleme is hiányzik működő baktérium ostorokból. Elég egyszerűen sikerült cáfolni ezt az érvelést. Hogy ezt az ellentmondást feloldja, William Dembski kicsit módosította az eredeti meghatározást:
"A system performing a given basic function is irreducibly complex if it includes a set of well-matched, mutually interacting, nonarbitrarily individuated parts such that each part in the set is indispensable to maintaining the system's basic, and therefore original, function. The set of these indispensable parts is known as the irreducible core of the system."
"Egy adott alapvető működést ellátó rendszer egyszerűsíthetetlenül összetett, ha tartalmaz egy készlet egymáshoz illő, egymással kölcsönható, nem tetszőlegesen kiegyénült részegységet, amely készlet minden eleme elengedhetetlen a rendszer alapvető, ezáltal eredeti működéséhez. Ezen elengedhetetlen részegységek a rendszer egyszerűsíthetetlenül összetett magját alkotják."
No most ez a meghatározás, attól eltekintve, hogy jóval ködösebb, mint Behe változata, ebben az esetben csak növeli a homályt, bár tény, hogy jóval megengedőbb, hiszen e maghatározás szerint nem kell az egész rendszernek egyszerűsíthetetlennek lennie, csak egy egyszerűsíthetetlen "magot" kell tartalmaznia. Mondjuk ez még mindig nem magyarázza meg, hogy akkor tulajdonképpen mit is tervezett az értelmes tervező? Mert ugye az E. coli baktérium ostorának az egyszerűsíthetetlen magja nem működik önmagában, kellenek hozzá olyan alkatrészek, amelyek más rendszerekből hiányoznak, tehát nyilván nem egyszerűsíthetetlenül összetettek. Viszont akkor fel kell tételeznünk, hogy az értelmes tervező egy működésképtelen rendszert alkotott, amit evolúciós folyamatok tettek működőképessé. Nem tudom, ez mennyire "értelmes" tervezés.
Létezik egy harmadik lehetőség is, nevezetesen az eredetileg tervezett ostor egy egyszerűbb, ámde működőképes szerkezet volt, amelyhez később kapcsolódtak újabb részegységek, amelyek részben átvették egyes eredeti alkatrészek működését, így ha egy mai ostorból eltávolítjuk őket, az már nem működik nélkülük, de ettől még későbbi toldalékok. Ez a magyarázat tetszetős, ugyanis éppen arra világít rá, hogyan alakulhat ki egy egyszerűsíthetetlenül összetett rendszer lépésenként, evolúciós folyamatok által, ebben az esetben az értelmes tervező már csak fölösleges csökevény az elméletben, ha eltávolítjuk, ugyanúgy megmagyarázza ezen rendszerek létrejöttét.
Ezen kívül is vannak gyenge pontok az elméletben: Az első gond vele, hogy ez esetben egy Gram-pozitív baktérium esetében egy másik rendszerről van szó, mint az E. coli esetében, amely tényt az elmélet egyszerűen nem kezeli sehogyan sem. Ezek szerint "a baktérium ostor" tulajdonképpen nagyon sok különböző, ám egytől-egyig egyszerűsíthetetlenül összetett rendszer gyűjtőneve. Viszont ezt a kérdést én még nem hallottam egyetlen ÉRTEM hívőtől se, illetve nem olvastam egyetlen írásukban sem, ők állandóan "a" baktérium ostorról beszélnek, ami alatt ki tudja hány különböző rendszert értenek, de valamiért kísérletet sem tesznek arra, hogy ezt a bizonytalanságot eloszlassák. Gyakorlatilag kizárólag az Escherichia coli baktérium ostoráról beszélnek, teljes mértékben figyelmen kívül hagyva a tényt, hogy ezen kívül nagyon sok ettől különböző baktériumostor létezik, amelyek felépítése romba dönti az elméletüket.
A másik érdekes része ennek a meghatározásnak, hogy e szerint a rendszer alapvető működése az eredeti feladata is, viszont ez a körkörös érvelés szép példája, hiszen eleve kizárja az esetleges evolúciós változásokat, azaz, amely rendszer egy más működést végző rendszerből alakult ki, máris nem egyszerűsíthetetlenül összetett.
Ha megvizsgáljuk közelebbről ezeket a "részegységeket", amelyekből ez az "egyszerűsíthetetlenül összetett" rendszer áll, megint csak meglepetésben lesz részünk. Mik is ezek a részegységek, amik elengedhetetlenek a működéséhez? Behe professzor biokémikus, ő az egyes fehérjéket tekinti részegységnek. Kiválasztottam hát két fehérjét hasraütéses alapon, egyet a táblázat elejéről, egyet a végéről, legyen mondjuk a FlgD és a MotA.
Mindkettő szándékosan olyan fehérje, amely minden eddig ismert ostorban megtalálható, így mindenképpen egyaránt bele kell hogy essen mindkét "egyszerűsíthetetlen összetettség" meghatározásba. Azt nyilván tudjuk, hogy a fehérjék aminosavakból állnak, így nézzük meg, mi ezen fehérjék aminosavsorrendje egyes élőlényekben!
Hasraütéses alapon kiválasztottam tíz FlgD fehérjét, kijelöltem őket a Uniprot adatbázisában (www.uniprot.org) és megpróbáltam az adatbázis beépített programjának a segítségével egymáshoz illeszteni őket. Az eredmény, azt hiszem megdöbbentő: ezek a fehérjék szinte nem is hasonlítanak egymásra.
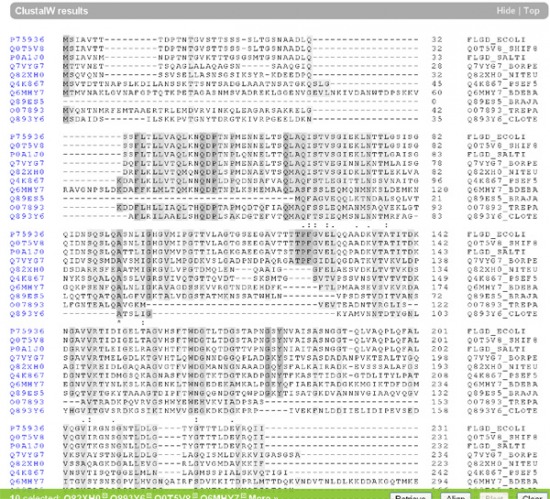
Ha egy megfelelő programmal számszerűsítjük is a különbséget, meglepő eredményeket kapunk:
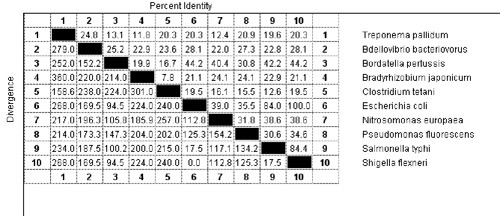
Ha ugyanígy megvizsgáljuk ugyanezen élőlények MotA fehérjéit, hasonló eredményt kapunk:
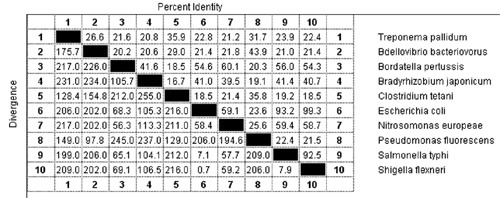
No most ezek a fehérjék egy "egyszerűsíthetetlenül összetett" rendszer alapelemei, amelyek az ÉRTEM elmélet szerint nem jöhettek létre evolúciós folyamatok során, hiszen ezek változásai működésképtelenné tennék az egész rendszert. Ezzel szemben azt látjuk, hogy maguk az alkotóelemek csak mérsékelt hasonlóságot mutatnak. Ha bárki szeretne, nyugodtan keresgélhet még a Uniprot adatai közt, nyilván hasonló képet lát majd. Az ÉRTEM hívei gyakran szoktak értelmes szöveggel példálózni, hogy alátámasszák mondanivalójukat. Hogy érzékeltessem, mit is jelent ez a különbség, fogtam egy ismert szöveget és kicseréltem benne egyes betűket. Elsőként a szöveg betűinek 22% -át hagytam változatlanul, a táblázatban több, hasonló mértékig azonos szekvencia is található, amelyek mind egy-egy működő baktérium ostor részei. Megkérlek, kedves olvasó, találd ki, mi ez a vers!
Eg, ft a hz! tdfgnft, krtd
A rthzests ldrtz gze bgzn?
Ldr, ftre gz he hujmn, hts gd,
Hjkl fgzeacvb h jene ghjklz!
Ugye nem ment! Lássuk, mit mondasz, ha a betűk felét cserélem ki!
Tj, ms a kd! tfghewsó, kgnd
A fzhbjbkn lalil izb brzt?
Lnk, cgjk jr ad iegen, hkt ád,
Hkly fdbvisre a keed dotkwt!
Még mindig elég zavaros. Ha mondjuk csak a betűk húsz százalékát cserélem ki, talán érthetőbb!
Eg, mr a kő! ttúkavyó, kend
A seobtbhn lhkik itt bhnt?
Lkm, ceak jh at ijten, jót ád,
Hohj föuvutte a kind dolgát!
Azért ebből már rá lehet jönni. A megfejtés: Petőfi Sándor: Anyám tyúkja. A feladat nyilván jóval könnyebb, mint egy ismeretlen szöveg megfejtése, mivel az iskolában mindannyiunknak meg kell tanulni ezt a verset, nyilván emlékszünk rá, valamint a szöveg tagolásán, az írásjelek elhelyezkedésén nem változtattam. Mindannyian ismerjük ezt a szövegrészt, mégis teljesen elveszik az információtartalma, ha a szöveget alkotó betűk nyolcvan százalékát lecseréljük. Ellenben ezek az FlgD fehérjék működő baktérium ostorok részei, azaz különbözőségük dacára mégis működnek, annak ellenére, hogy egymásra csak olyan húsz százalékban hasonlítanak. Itt kezdenek szaporodni a kínos kérdések: Tulajdonképpen melyik FlgD fehérjét teremtette az értelmes tervező? Az ÉRTEM hívei valamiért az E. coli baktérium ostorát tűzték zászlajukra, de igazából arra semmilyen magyarázatot sem adnak, hogy miért éppen ezt tartják az értelmes tervező keze munkájának. Honnan tudjuk, hogy éppen az volt az "eredeti", nem egy másik? Miért nem éppen a Bdellovibrio bacteriovorus ostorát tervezte az értelmes tervező? Hogyan jött létre a többi? Mert ugye ha evolúciós folyamatok során, mutációkkal-szelekcióval, akkor tulajdonképpen az "egyszerűsíthetetlenül összetett" baktérium ostor egy adott alkatrészének a nyolctizedét evolúciós folyamatok alakították ki. Miből gondoljuk, hogy a többit nem? Ha az egyes alkatrészek ilyen mérvű változásokat képesek elviselni és a rendszer mégis működőképes marad, miért tűnik olyan lehetetlennek, hogy eleve ezek a változások alakították ki? Ha pedig feltételezzük, hogy az értelmes tervező tervezte ezt a rengeteg különféle ostort, minek fáradt ennyit? Nekem valahogyan túl sok fölösleges munkának tűnik FlgD fehérjék ezreit, tízezreit megtervezgetni, de ezzel az ÉRTEM hívei is így vannak. Ha emlékszünk még: "Egy intelligens cselekvő számára logikus és egyszersmind hatékony is az élőlényeket közös biokémiai alapon megtervezni." (http://ertem.hu/content/view/173/34/) Viszont akkor miért nem így cselekedett a feltételezett értelmes tervező?
Nyilván nem tudjuk pontosan, minden kétséget kizáróan bizonyíthatóan megmondani, hogyan alakult ki a baktérium ostora, mert olyan élőlényekről van szó, akikből kövületek nem maradnak fenn, ráadásul a vizsgálni kívánt folyamat valamikor három- négymilliárd éve történt, az akkori köztes alakok nyilván azóta rég kihaltak. Akármilyen részletes modellt vázolnánk is fel, meg lehetne kérdezni, hogy jó, jó, szép, szép, akár így is történhetett volna, de mi bizonyítja, hogy tényleg így történt? A válasz nyilván az, hogy ezt sajnos nem tudjuk bizonyítani, ez csak egy lehetséges modell lenne. Viszont ha feltételezzük, hogy evolúciós folyamatok alakították ki, abból következik hogy kellettek lenni egyszerűbb felépítésű ostoroknak is, Behe professzor elmélete pedig bevallottan arra épül, hogy ilyenek nem létezhettek. Viszont mivel ilyen egyszerűbb ostorokat sikerült találni, Behe professzor elmélete hibás, az elméletből következő előrejelzéseket maga a valóság cáfolja.
A fenti gyors körképből egyértelműen kitűnik, hogy "a baktérium ostor" nem egyszerűsíthetetlenül összetett rendszer. Ha megvizsgáljuk Behe professzor többi példáját, ugyanezt tapasztaljuk:
A véralvadási kaszkád fehérjéi is az egyszerűsíthetetlenül összetett rendszerek közé tartoznak Behe professzor szerint, ám 1998 –ban, két évvel a Darwin fekete doboza megjelenése után kiderült, hogy a bálnákból hiányzik a XII –es véralvadási faktor. A génje megtalálható, ám két mutáció működésképtelené tette, pszeudogén (Semba U. és munkatársai 1998.). 2003 –ban pedig kiderült, hogy a gömbhal (Fugu rubripes) véralvadási kaszkádjából öt fehérje hiányzik, amelyek az emlősökben megvannak (Jiang Y és munkatársai 2003.).
Az adaptív immunitást is ide sorolta Behe professzor az eredeti könyvében, ám azóta többen is tanulmányozták az állkapocs nélküli gerincesek immunrendszerét, és meglepő különbségeket fedeztek fel: Ezekben az élőlényekben is véletlenszerű szomatikus rekombinációval alakulnak ki az immunsejtek receptorai, ám különbözik a felhasznált szignálszekvencia, nyoma sincs a magát a rekombinációt katalizáló az állkapcsos gerincesekben meglévő RAG1 és RAG2 enzimeknek, valamint a receptorok teljesen különböző szerkezetűek, az állkapcsos gerincesek immunglobulin doméneket tartalmazó receptora helyett az állkapocsnélküli gerincesekben leucin- gazdag repeat doméneket tartalmaznak a szomatikus rekombinációval előálló receptorok. (Guo P. és munkatársai 2009.)
Az eukarióták csillói is Behe eredeti példái között voltak, sajnos ezekből is ismert egyszerűbb változat. (Woolley D. M. 1997.)
Az elektronszállításról már írtam, ez sem egyszerűsíthetetlenül összetett.
A telomérák fenntartása is egyszerűsíthetetlenül összetett rendszer Behe professzor szerint, nagy kár, hogy az ecetmuslicában például nem az emlősökben megismert telomeráz enzimen alapuló rendszer tartja fenn a kromoszómák végein található szakaszokat, hanem egy módosult, megszelídített ugráló genetikai elem. (Capkova Frydrychova R. és munkatársai 2008.)
Lényeg, ami lényeg, ha egy egyszerűsíthetetlenül összetett rendszerből létezik egyszerűbb változat, az önmagában agyoncsapja a rendszer egyszerűsíthetetlenségét és bizony az erre felhozott példák mindegyikéről kiderült időközben, hogy bizony léteznek egyszerűbb változataik is, azaz nem egyszerűsíthetetlenül összetettek. Hogy újra idézzem az ÉRTEM hivatalos honlapját: "Egyszerűsíthetetlen összetettség nem állhat elő fokozatosan, egy korábbi rendszer jelentéktelen, egymás utáni változásainak eredményeként, mert definíció szerint egy egyszerűsíthetetlenül összetett rendszernek bármely őse működésképtelen." Erre mondják, hogy ipi-apacs. Az úgynevezett "egyszerűsíthetetlenül összetett" rendszerek nem azok és kész.
Egyetlen kérdést nem érintettünk csak eddig, azt a tényt, amelyet sehol sem fektettek le írásban. Annak idején leveleztem az ÉRTEM hazai apostolaival, akkor döbbentem rá erre a kimondatlan törvényre. Ha bárki más rendszert próbál példaként állítani, mint amelyeket Behe professzor említett annak idején a könyvében, rögtön megmagyarázták, hogy ez csak egy szokásos evolucionista csalás, egy nyilvánvalóan nem-egyszerűsíthetetlenül összetett rendszert veszünk és arról bizonyítjuk, hogy nem is egyszerűsíthetetlenül összetett. Ekkor jöttem rá, hogy az egyszerűsíthetetlenül összetettségnek létezik egy mindennél fontosabb követelménye is, amelyet azonban sohasem foglaltak írásba: Az adott rendszert az ÉRTEM apostolai ismerjék el annak.
Mondanék egy nagyon egyszerű példát, az ember szaporodását. Ugye mi méhlepényes emlősök vagyunk, az utódaink az anya méhében fejlődnek kilenc hónapig, csak ez után születnek meg, az újszülötteket ez után még hosszú ideig az anya tejével táplálja. Ez a rendszer, vagyis az ember szaporodása az egyszerűsíthetetlenül összetett rendszer minden követelményét kielégíti: Egységes rendszer, számos egymással kölcsönható elemből áll (petefészkek, méh, méhlepény, tejmirigyek, az egész folyamatot szabályozó hormonok, ezek receptorfehérjéi), amelyek mindegyike hozzájárul az alapvető működéshez és amelyek közül bármelyik eltávolítása működésképtelenné teszi az egész rendszert. Ezen kívül számos előnye van még ennek a példának: Gyakorlatilag mindenki ismeri valamennyire a működését, hiszen egy bizonyos koron túl már mindenki tisztában van azzal, hogyan is lesznek a kisbabák. Eléggé jól kutatott rendszer, számos emlősállat szaporodását ismerjük részletesen, nyilván számos példát lehetne rá hozni. Ráadásul itt a rendszer minden egyes elődjének működőképesnek kell lennie, hiszen egy szaporodásképtelen állat utódok híján nyilvánvalóan nem lehet a mai emlősök őse.
Az evolúció elmélete feltételezi, hogy ez a bonyolult, minden jel szerint egyszerűsíthetetlenül összetett rendszer a hüllők szaporodásából fejlődik ki, akik csak raknak egy tojást és kész. Ugyanúgy elő lehetne hozni az érveket, miszerint az ember szaporító szervrendszerének nem lehettek működőképes elődei, hiszen bármelyik elemének eltávolítása az egész rendszert működésképtelenné teszi és ugyanígy fel lehetne hozni az összes szokásos érvet, majd levonni a következtetést, hogy az értelmes tervező tervezte. Igazán tetszetős lenne, nem? Mégiscsak sokkal de sokkal közérthetőbb lenne, mint a baktérium ostorral példálózni, amiről lássuk be, a legtöbben azt sem tudják eszik –e vagy isszák?
Azonban van egy apró bökkenő, nevezetesen az emlősök ősei nem haltak ki mind, a hüllők szaporító szervrendszere és az emlősök szaporító szervrendszere közti átmeneti alakok közül ma is sok él. A tojásrakó emlősök lágy héjú tojásokkal szaporodnak, de a kikelő utódaikat tejmirigyeik váladékával táplálják. Az erszényes emlősök utódai méhlepény hiányában csak rövid ideig fejlődhetnek az anyaméhben, így igen fejletlen állapotban kell megszületniük, az anyaállat erszényében fejlődnek aztán önállóan is életképes utódokká. Vagyis egy ilyen bonyolult egyszerűsíthetetlenül összetett rendszer is kialakulhat lépésenként, apróbb változtatásokkal, egyszerűbb ősökből. Akkor tulajdonképpen miért gondoljuk, hogy a többi nem? Ugye lehet azzal érvelni, hogy az ember szaporítószervrendszere nem egyszerűsíthetetlenül összetett, ez csak egy hajánál fogva előrángatott példa, amit szándékosan hoztam. Viszont akkor jó volna megmagyarázni, hogy miért nem az? Van esetleg ezeknek a meghatározásoknak egy titkos záradéka, amit kívülállók nem ismerhetnek meg, csak Behe professzor? Erre a rendszerre az eredeti és a bővített meghatározások mindegyike ráillik, egyetlen "bűne" van, hogy tudjuk, hogyan alakult ki, mivel az ősi, egyszerűbb rendszerek közül több fennmaradt a mai napig. Ha teszem azt már kihaltak volna a tojásrakó emlősök és az erszényesek, vajon ez is egyszerűsíthetetlenül összetett rendszer lenne? A személyes véleményem az, hogy az a bizonyos titkos záradék egyszerűen annyi, hogy csak az a rendszer egyszerűsíthetetlenül összetett, amelyről nem tudjuk pontosan, hogyan alakult ki. Behe eredeti meghatározása számos rendszerre tökéletesen illik, amelyek kialakulását ismerjük, ezek azonban nem számítanak valódi egyszerűsíthetetlenül összetett rendszernek, hiszen tudjuk, hogyan alakultak ki. Nem mellesleg ez a körkörös érvelés szép példája: Nem tudjuk hogyan alakultak ki az egyszerűsíthetetlenül összetett rendszerek, hiszen csak azt ismerjük el annak, amiről nem tudni, hogy alakult ki, aminek a kialakulása ismert, azok nem egyszerűsíthetetlenül összetettek. Amennyiben nincs igazam, felkérném a kritikusokat, hogy magyarázzák meg, miért nem egyszerűsíthetetlenül összetett az ember szaporítószervrendszere, vagy bármely más olyan rendszer, amelyre tökéletesen ráillik Behe professzor meghatározása, de mégsem fogadják el annak! "Az egyszerűsíthetetlen összetettségre valójában egy biokémikus tankönyv valamennyi oldalán találhatunk példát." (http://ertem.hu/content/view/31/16/)
Összefoglalva Behe professzor sokszor varázsszóként emlegetett "egyszerűsíthetetlenül összetett" rendszerei tulajdonképpen a rések istene ("god of the gaps") érvelés újabb köntösben. A tudásunknak mindig lesznek házagai, ha úgy tetszik határai. Mindig lesznek folyamatok, amelyeket nem ismerünk teljes részletességgel, vagy egyáltalán nem, mert eddig még nem kutatta senki. Kijelenthetjük, hogy ott lakik Isten, de annál kínosabb, ha a tudásunk határai kicsit kijjebb kerülnek, például új felfedezések nyomán és kiderül, hogy mégsem. Behe professzor minden példaként hozott egyszerűsíthetetlenül összetett rendszeréből ismerünk egyszerűbb, ám működő változatokat, az elmélete egyszerűen megdőlt. Ezek után meglepő, hogy mégis tizenhárom éve töretlen lendülettel véres kardként hordozzák körbe az ÉRTEM hívei, mint a hitük fő bizonyítékát, annak ellenére, hogy az elméletet gyakorlatilag megjelenésének pillanatában meg is cáfolták biológusok, ráadásul az azóta napvilágra került tények újra és újra csak Behe professzor elméletét cáfolják.
Capkova Frydrychova R, Biessmann H, Mason J M. (2008): Regulation of telomere length in Drosophila; Cytogenetic and Genome Research 122(3-4):356-64
Guo P, Hirano M, Herrin BR, Li J, Yu C, Sadlonova A, Cooper MD (2009): Dual nature of the adaptive immune system in lampreys; Nature 459(7248):796-801
Hizukuri Y., Yakushi T., Kawagishi I., Homma M. (2006): Role of the intramolecular disulfide bond in FlgI, the flagellar P-ring component of Escherichia coli; Journal of Bacteriology 188(12):4190-7
Jiang Y, Doolittle RF. (2003): The evolution of vertebrate blood coagulation as viewed from a comparison of puffer fish and sea squirt genomes; Proc Natl Acad Sci U S A. 100(13):7527-32.
Pallen M. J., Matzke N. J. (2006): From The Origin of Species to the origin of bacterial flagella; Nature Reviews Microbiology 4, 784-790
Semba U, Shibuya Y, Okabe H, Yamamoto T. (1998): Whale Hageman factor (factor XII): prevented production due to pseudogene conversion; Thrombosis Research 90(1):31-7.
Woolley D. M. (1997): Studies on the eel sperm flagellum. I. The structure of the inner dynein arm complex; Journal of Cell Science. 110 ( Pt 1):85-94
Ha az ÉRTEM oldaláról érkeztél ide, kérlek olvasd el a cikksorozat többi részét is!
Nem értik 1. - A nejlonevő baktérium
Nem értik 2. - A polip szeme
Értelmes tervezés - A P-elem
Nem értik 3. - Genetika és makroevolúció
A mutációkról
A fajok eredete
Biokémiai hasonlóságok
Nem értik 5. - A specifikus összetettség
Nem értik 6. - Az értelmes tervezés mozgalom tudományos tevékenysége
 (Vendégmunkásunk, SexComb, újra lesújt.)
(Vendégmunkásunk, SexComb, újra lesújt.)
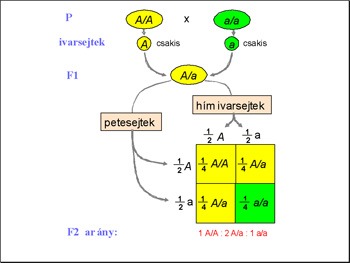 Ezek után az F1 nemzedék
egyedeit egymással keresztezte és meglepődve tapasztalta, hogy a
körükben ismét megjelentek fehér virágú egyedek is. Mendel lángelméje
ott mutatkozott meg, hogy meg is számolta az utódokat, azaz statisztikai
alapon elemezte őket! 705 darab piros virágú és 224 darab fehér
virágú növényt kapott. Ezt az arányt ő 3:1 –nek mérte. Ezek
után kíváncsi volt rá, hogy az utódok vajon tiszta vonalúak –e?
Az F2 nemzedék egyedeit önmegtermékenyítéssel szaporította és
megvizsgálta az utódaikat (F3 nemzedék). Azt tapasztalta, hogy a
domináns fenotípust mutató egyedek harmada tiszta vonalú, azaz ebben
az esetben az utódaik is piros virágúak lesznek, míg a kétharmaduk
nem tiszta vonalú, azaz az utódaik között fehér virágúak is felbukkannak,
míg a fehér virágúak mind tiszta vonalúak. Arra következtetett,
hogy a piros virágúak egyharmada, azaz az összes utód egynegyede
AA genotípusú, azaz tiszta vonalú, kétharmada, azaz az összes utód
fele Aa genotípusú, azaz fenotípusosan piros virágú, ám az utódaik
között fehér virágúak is megjelennek, míg a fehér virágúak
aa genotípusúak, természetesen tiszta vonalúak.
Ezek után az F1 nemzedék
egyedeit egymással keresztezte és meglepődve tapasztalta, hogy a
körükben ismét megjelentek fehér virágú egyedek is. Mendel lángelméje
ott mutatkozott meg, hogy meg is számolta az utódokat, azaz statisztikai
alapon elemezte őket! 705 darab piros virágú és 224 darab fehér
virágú növényt kapott. Ezt az arányt ő 3:1 –nek mérte. Ezek
után kíváncsi volt rá, hogy az utódok vajon tiszta vonalúak –e?
Az F2 nemzedék egyedeit önmegtermékenyítéssel szaporította és
megvizsgálta az utódaikat (F3 nemzedék). Azt tapasztalta, hogy a
domináns fenotípust mutató egyedek harmada tiszta vonalú, azaz ebben
az esetben az utódaik is piros virágúak lesznek, míg a kétharmaduk
nem tiszta vonalú, azaz az utódaik között fehér virágúak is felbukkannak,
míg a fehér virágúak mind tiszta vonalúak. Arra következtetett,
hogy a piros virágúak egyharmada, azaz az összes utód egynegyede
AA genotípusú, azaz tiszta vonalú, kétharmada, azaz az összes utód
fele Aa genotípusú, azaz fenotípusosan piros virágú, ám az utódaik
között fehér virágúak is megjelennek, míg a fehér virágúak
aa genotípusúak, természetesen tiszta vonalúak.
 Nézzünk meg inkább egy példát,
ahol a háziasítás folyamata jól követhető sőt, még az egyes
tulajdonságok öröklődése is ismert. Gondolom nem kell bemutatnom
a szíriai aranyhörcsögöt, amely kedvelt háziállat nálunk is.
Latin neve Mesocricetus auratus. Magyar nyelven is bőséges
forrásanyag áll a rendelkezésünkre róla (pl.
Nézzünk meg inkább egy példát,
ahol a háziasítás folyamata jól követhető sőt, még az egyes
tulajdonságok öröklődése is ismert. Gondolom nem kell bemutatnom
a szíriai aranyhörcsögöt, amely kedvelt háziállat nálunk is.
Latin neve Mesocricetus auratus. Magyar nyelven is bőséges
forrásanyag áll a rendelkezésünkre róla (pl.  (Sorozatunk
(Sorozatunk 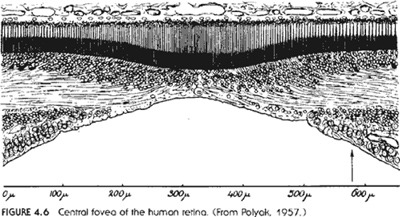 Ezen a
területen ugyanis az idegsejtek mintegy "elhúzódnak"
a fényérzékelő sejtek fölül, így
téve lehetővé az éles látást.
Itt azért felmerül a kérdés, hogy az emberi
szem miért inverz szem, ha azon a ponton, ahol az éles
kép keletkezik, az idegsejtek kénytelenek elhajolni a
fényérzékeny sejtek fölül? Az
idegsejtek léte valóban szükséges a szem
működéséhez, ők továbbítják
a fényérzékeny sejtekben keletkező
ingerületet, de akkor miért éppen arról a
területről húzódnak el (természetesen
nem hiányoznak, a látógödör körüli
idegsejtek nyúlványai továbbítják
az itt található fényérzékeny
sejtek ingerületét), ahol a legjobban kellenének,
hiszen nyilván itt a legnagyobb az igény az
adatfeldolgozásra és a legtöbb ingerület is
itt keletkezik. Milyen magyarázatot találtak az
értelmes tervezés hívei a sárgafolt
felépítésére? Miért kell az
idegsejteknek szabadon hagyni a fényérzékeny
sejteket éppen azon a ponton, ahol éles képet és
színeket is látunk, ha az emberi szem azért
inverz szem, mert másképpen nem adhatna ilyen minőségű
képet? Mi ennek az ésszerű indoka? A boncolgatott
cikkben miért nem említik meg az emberi szemnek ezt a
meglehetősen fontos jellegzetességét, ha már
a polip szemének hasonló területét
részletesen tárgyalják? Első pillantásra
látszik, hogy ez egy értelmesen tervezett szerv?
Ezen a
területen ugyanis az idegsejtek mintegy "elhúzódnak"
a fényérzékelő sejtek fölül, így
téve lehetővé az éles látást.
Itt azért felmerül a kérdés, hogy az emberi
szem miért inverz szem, ha azon a ponton, ahol az éles
kép keletkezik, az idegsejtek kénytelenek elhajolni a
fényérzékeny sejtek fölül? Az
idegsejtek léte valóban szükséges a szem
működéséhez, ők továbbítják
a fényérzékeny sejtekben keletkező
ingerületet, de akkor miért éppen arról a
területről húzódnak el (természetesen
nem hiányoznak, a látógödör körüli
idegsejtek nyúlványai továbbítják
az itt található fényérzékeny
sejtek ingerületét), ahol a legjobban kellenének,
hiszen nyilván itt a legnagyobb az igény az
adatfeldolgozásra és a legtöbb ingerület is
itt keletkezik. Milyen magyarázatot találtak az
értelmes tervezés hívei a sárgafolt
felépítésére? Miért kell az
idegsejteknek szabadon hagyni a fényérzékeny
sejteket éppen azon a ponton, ahol éles képet és
színeket is látunk, ha az emberi szem azért
inverz szem, mert másképpen nem adhatna ilyen minőségű
képet? Mi ennek az ésszerű indoka? A boncolgatott
cikkben miért nem említik meg az emberi szemnek ezt a
meglehetősen fontos jellegzetességét, ha már
a polip szemének hasonló területét
részletesen tárgyalják? Első pillantásra
látszik, hogy ez egy értelmesen tervezett szerv?
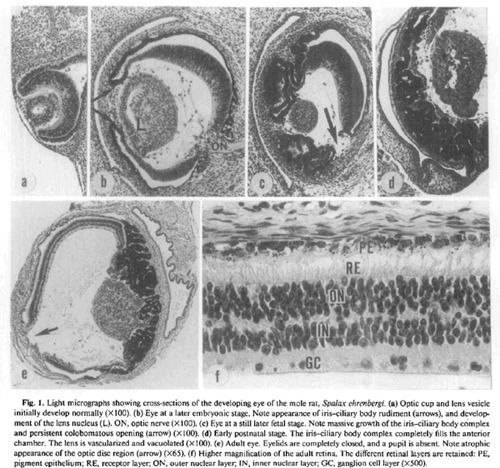 Sanyal S.
és munkatársai 1990. íródott cikkében
szemléletes ábra található a földikutya
szemének fejlődéséről. A szem a látó
emlősökéhez hasonlóan kezd fejlődni, a
korai szem (a) minden szempontból egy tíznapos
egérembrióéhoz hasonló. Megjelenik a
szemlencse (L), és a (b) ábrán láthatóak
a szivárványhártya és a sugártest
kezdeményei is (nyilak). Majd ezek után az embrió
szemének fejlődése teljesen más irányt
vesz: a szemlencse mérete lecsökken, gyakran vérerek
nyomulnak bele, átlátszatlanná válik, a
szem mérete messze nem éri el a hasonló
testtömegű, látó gerincesek szemméretét,
a fejlődésnek indult szemlencse a szemfenékre
tapad, ezzel a más gerincesek szemében betöltött szerepére alkalmatlanná válik. Az (f) kép a földikutya retináját
mutatja, amelyben tisztán felismerhető a látó
emlősök szemére jellemző szerveződés.
PE – az annyit emlegetett pigmentréteg; RE –
fényérzékelő sejtek; ON, IN GC – a
retina idegsejtjei. Ha megfigyeljük, a földikutya szeme is
inverz szem, mint az összes többi emlősé, itt
is tisztán felismerhetőek a fényérzékeny
sejtek és a beérkező fény között
található idegsejtek. Hogy kicsit visszautaljak a cikk
elejére: neki nem teljesen mindegy? Maga az állat vak,
az ő fényérzékeny sejtjei akárhogyan
állhatnának, mégis tisztán felismerhető
az összes többi emlős szemére is jellemző
szerkezet. Ennek a magyarázata az evolúció
talaján maradva egyszerű: egy, a többi emlőséhez
hasonló szem elcsökevényesedésével
alakult ki ez a fényérzékelő szerv, így
bár eredeti feladatát már nem tölti be,
alapszerkezete mégis tisztán felismerhető. Mint korábban már említettem, itt
éppen megfigyelhető egy új szerv kialakulása
egy már meglévőből. Mi az értelmes
tervezés talaján nyugvó magyarázat? Első
pillantásra látszik hogy ez egy értelmesen
tervezett szerv? Ez egy egyszerűsíthetetlenül
összetett szerv? Például miért alakul ki a
szemlencse, ha láthatóan nem működik? Miért
vannak meg a színlátás elemei is ebben a
csökevényes szemben, ha csak a fény erősségét
érzékeli? Miért azonos a földikutya és
az ember retinájának alapszerkezete? (Szerk. megj: részben hasonló szem-sorvadás zajlik
Sanyal S.
és munkatársai 1990. íródott cikkében
szemléletes ábra található a földikutya
szemének fejlődéséről. A szem a látó
emlősökéhez hasonlóan kezd fejlődni, a
korai szem (a) minden szempontból egy tíznapos
egérembrióéhoz hasonló. Megjelenik a
szemlencse (L), és a (b) ábrán láthatóak
a szivárványhártya és a sugártest
kezdeményei is (nyilak). Majd ezek után az embrió
szemének fejlődése teljesen más irányt
vesz: a szemlencse mérete lecsökken, gyakran vérerek
nyomulnak bele, átlátszatlanná válik, a
szem mérete messze nem éri el a hasonló
testtömegű, látó gerincesek szemméretét,
a fejlődésnek indult szemlencse a szemfenékre
tapad, ezzel a más gerincesek szemében betöltött szerepére alkalmatlanná válik. Az (f) kép a földikutya retináját
mutatja, amelyben tisztán felismerhető a látó
emlősök szemére jellemző szerveződés.
PE – az annyit emlegetett pigmentréteg; RE –
fényérzékelő sejtek; ON, IN GC – a
retina idegsejtjei. Ha megfigyeljük, a földikutya szeme is
inverz szem, mint az összes többi emlősé, itt
is tisztán felismerhetőek a fényérzékeny
sejtek és a beérkező fény között
található idegsejtek. Hogy kicsit visszautaljak a cikk
elejére: neki nem teljesen mindegy? Maga az állat vak,
az ő fényérzékeny sejtjei akárhogyan
állhatnának, mégis tisztán felismerhető
az összes többi emlős szemére is jellemző
szerkezet. Ennek a magyarázata az evolúció
talaján maradva egyszerű: egy, a többi emlőséhez
hasonló szem elcsökevényesedésével
alakult ki ez a fényérzékelő szerv, így
bár eredeti feladatát már nem tölti be,
alapszerkezete mégis tisztán felismerhető. Mint korábban már említettem, itt
éppen megfigyelhető egy új szerv kialakulása
egy már meglévőből. Mi az értelmes
tervezés talaján nyugvó magyarázat? Első
pillantásra látszik hogy ez egy értelmesen
tervezett szerv? Ez egy egyszerűsíthetetlenül
összetett szerv? Például miért alakul ki a
szemlencse, ha láthatóan nem működik? Miért
vannak meg a színlátás elemei is ebben a
csökevényes szemben, ha csak a fény erősségét
érzékeli? Miért azonos a földikutya és
az ember retinájának alapszerkezete? (Szerk. megj: részben hasonló szem-sorvadás zajlik 
Utolsó kommentek