
A rovarok körében általában a két pár szárny dívik (gondoljunk csak a szitakötőkre vagy a bogarakra), amelyek a második ill. harmadik tor szelvényről erednek. Akad azonban egy rend, amely csak egyetlen pár hártyás szárnnyal rendelkezik, ez a Kétszárnyúak rendje, a Diptera. Ezekben a rovarokban a második pár szárny egy különleges egyensúlyozó szervvé, ún. billérré alakult - mégpedig, mint azt mindenki, aki valaha kergetett legyet vagy szúnyogot tudja, elég sikeresen...
Már hosszú ideje tudott, hogy a változás mögött egy homebox gén áll, az Ultrabithorax (Ubx), ui. mai napig a hometikus mutációk prototípusa az ebben a génben bekövetkezett bithorax mutáció, amelynek eredményeként a muslica billérjei helyett is szárnyak fejlődnek. Normális esetben ennek a homeobox génnek a terméke jelen van a billért hordozó harmadik tor szelvényben, és hiányzik a szárnyat hordozó másodikból, de hogy az Ubx azonban pontosan hogyan is fejti ki a hatását, az mindmáig homályban maradt.

A bal oldalon egy vad típusú (azaz normális) Drosophila látható, míg jobb oldalon egy Ubx mutáns.
 Most azonban a Science egyik cikke fellebbenti a fátylat (legalább is részben) az Ubx hatásmechanizmusának mikéntjéről. A szárnyak méretét a Decapentaplegic (Dpp) fehérje szabályozza - ha több van belőle, jobban osztódnak a sejtek - s a kísérletek tanúsága szerint ezen fehérje szabályozása változott meg a billérek kialakulása során.
Most azonban a Science egyik cikke fellebbenti a fátylat (legalább is részben) az Ubx hatásmechanizmusának mikéntjéről. A szárnyak méretét a Decapentaplegic (Dpp) fehérje szabályozza - ha több van belőle, jobban osztódnak a sejtek - s a kísérletek tanúsága szerint ezen fehérje szabályozása változott meg a billérek kialakulása során.
A muslica lárva fejlődése során megjelenő szárny (wing - W) kezdemények egyáltalán nem hasonlítanak a kifejlett szárnyakra, inkább miniatűr nyalókáknak tűnnek, amelyek csak a fejlődés egy későbbi fázisában laposodnak el. Ennek a "nyalókának" a hossztengelyében - amely a későbbi szárny élének felel meg - fejeződik ki a dpp gén, s innen diffundál a róla átíródó Dpp fehérje a környező sejtek közé. Mint arra a kutatók felfigyeltek, a billér (haltere - H) kezdeményében sokkal kevésbé expresszálódik a dpp és a Dpp aktivitás (ezt a kék színű P-Mad jelöli) is kevesebb sejtben figyelhető meg. Sőt, hogy az ok-okozati összefüggést megerősítsék: a Dpp aktivitást felerősítve a billérben, annak mérete 40%-al megnőtt, a szárnyban csökkenő aktivitás pedig 30%-os méretbeli csökkenést okozott.
Mi lehet tehát a billérben Dpp aktivitás csökkenésnek az oka? Mind a szárnyban, mind a billérben a Dpp egyik receptora, a Thickveinless (Tvk) komplementer módon fejeződik ki a Dpp aktivitással. Ennek az az oka, hogy ahol a receptor jelen van, ott megköti a Dpp-t ezáltal mintegy kihalássza, kititrálja a sejtközi folyadékból, azaz a Dpp nem tud tovább diffundálni. Azonban míg a szárnyban a Tkv nincs (vagy alig van) jelen a Dpp-t kifejező sejtek környezetében, addig a billér kezdeményben a Tkv minden sejtben egyforma intenzitással jelen van - így aztán persze a Dpp aktivitás is kevesebb sejtben érzékelhető, hiszen hamar "elfogy" a fehérje a sejtek közül.
Az Ubx valószínűleg a Tkv egyik negatív szabályozója, a master of thickveinless (mtv) gén elnyomásával hozza létre (részben) a szárny és billér között észlelt különbséget: ez a gén igen magas szinten expresszálódik a szárny hossztengelyében, míg a billérében nem. Ezért aztán a szárny hossztengelyében kevés Tkv keletkezik, s a Dpp szabadabban terjedhet.
Slusszpoénként: a nagyon szemfüleseknek esetleg feltünhetett, hogy a Dpp hasonlóan szabályozza egy szerv méretét mint a BMP egyes gerincesekben (lásd itt és itt), ami nem is igazán meglepő, figyelembe véve, hogy a Dpp a BMP gerinctelen megfelelője - csak éppen a rovarok esetében erre a furcsa névre hallgat ;-)).
Crickmore, MA, Mann, RS (2006) Hox Control of Organ Size by Regulation of Morphogen Production and Mobility. Science 313: 63-68.
 Most német és brit kutatóknak a hímivarsejtek esetében sikerült tovább menniük egy lépéssel és funkcionális spermiumokat létrehozniuk. Az eljárás során retinol-savban (RA) növesztettek olyan embrionális őssejteket, amelyekbe bejuttatak egy, a stra8 gén promóteréhez kötött fluoreszcens fehérjét (EGFP) kódoló génszakaszt. (A stra8 gén jellemzően csak a hímivarsejt-vonalban fejeződik ki, így a promóteréhez kötött EGFP is csak azokban a sejtekben kerül átírásra, amelyek ezirányba kezdtek differenciálódni - értsd az ilyen sejtek megfelelő megvilágítás esetén a GFP miatt zölden fluoreszkálnak.) A GFP pozitív sejteket elkülönítették, és a biztonság miatt egy másik hímivarsejt specifikus gén, a prm1 promóterével össekapcsolt piros fluoreszcens fehérje (DsRed) DNS-ét is beléjük juttatták (bízva abban, hogy azok a sejtek, amelyek mind a zöld, mind a piros fluoreszcens fehérjét expresszálják valóban hímivarsejtek lesznek). Az egyaránt piros-zöld sejteket (a kék színű DAPI a sejtmagok vizualizálására használt) közelebről megnézve, azok valóban expresszálták a spermatogenezishez kapcsolódó géneket, vagyis minden arra mutatott, hogy ezek funkcionális ős-hímivarsejtek.
Most német és brit kutatóknak a hímivarsejtek esetében sikerült tovább menniük egy lépéssel és funkcionális spermiumokat létrehozniuk. Az eljárás során retinol-savban (RA) növesztettek olyan embrionális őssejteket, amelyekbe bejuttatak egy, a stra8 gén promóteréhez kötött fluoreszcens fehérjét (EGFP) kódoló génszakaszt. (A stra8 gén jellemzően csak a hímivarsejt-vonalban fejeződik ki, így a promóteréhez kötött EGFP is csak azokban a sejtekben kerül átírásra, amelyek ezirányba kezdtek differenciálódni - értsd az ilyen sejtek megfelelő megvilágítás esetén a GFP miatt zölden fluoreszkálnak.) A GFP pozitív sejteket elkülönítették, és a biztonság miatt egy másik hímivarsejt specifikus gén, a prm1 promóterével össekapcsolt piros fluoreszcens fehérje (DsRed) DNS-ét is beléjük juttatták (bízva abban, hogy azok a sejtek, amelyek mind a zöld, mind a piros fluoreszcens fehérjét expresszálják valóban hímivarsejtek lesznek). Az egyaránt piros-zöld sejteket (a kék színű DAPI a sejtmagok vizualizálására használt) közelebről megnézve, azok valóban expresszálták a spermatogenezishez kapcsolódó géneket, vagyis minden arra mutatott, hogy ezek funkcionális ős-hímivarsejtek.
 Bár élő dodót mintegy 300 éve senki sem látott, mégis ez a nagyra nőtt (és alapvetően ronda) galamb a természetvédelem ikonikus alakjává vált - talán nem kis részben azért, mert Gerald Durrell is őt választotta
Bár élő dodót mintegy 300 éve senki sem látott, mégis ez a nagyra nőtt (és alapvetően ronda) galamb a természetvédelem ikonikus alakjává vált - talán nem kis részben azért, mert Gerald Durrell is őt választotta  Ha már
Ha már  A jelek szerint a történelem valóban ismétli önmagát, különösen ha a szőrszín megváltozásáról van szó.
A jelek szerint a történelem valóban ismétli önmagát, különösen ha a szőrszín megváltozásáról van szó.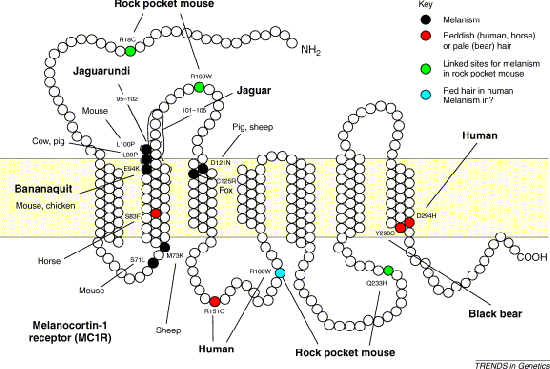
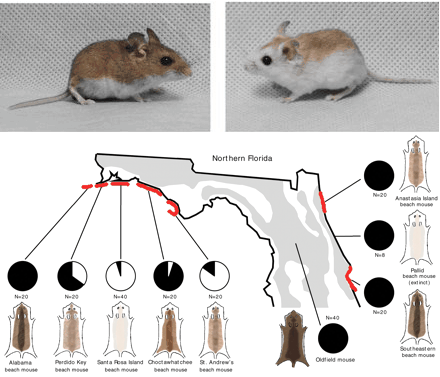

Utolsó kommentek