
 (primavis kollega vendégposztja következik)
(primavis kollega vendégposztja következik)
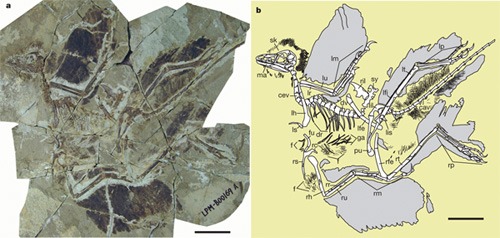
Az ÉK- Kinában található Liaoning tartomány tollas dinoszauru-szokról már világszerte ismert mezozoikumi lelőhelyei újabb „csodalényt” szolgáltattak. Ráadásul ez jóval idősebb is az eddigi leleteknél, hiszen korát 155-159 millió évre becsülik.
A majdnem komplett csontvázú, holló méretű s szinte teljesen tollakkal borított maradvány az Anchiornis huxleyi nevet kapta és a Liaoning Őslénytani Múzeum tulajdonát képezi.
Lelőhelye Daxishan településen található (Jianchang megye) és a Tiaojishan Formációhoz tartozik. Ez utóbbit a Középső-Jura – Felső-Jura határára helyezik, s a lelet is a Felső-Jura legalsó emeletének (Oxfordi) felel meg. Tehát valamennyivel idősebb a közismert bajorországi solnhofeni palarétegek koránál, amelyeket kb. 150 millió évre becsülnek (Tithoni emelet) s amelyekből az első Archaeopteryx leletek származnak.
Az új tollas lény rendkívüli hasonlatosságot mutat a nála jóval fiatalabb (125 millió éves) Microraptor-ral, elsősorban az alsó végtag tollakkal való borítottságát illetően.
Csonttani jellegzetességeit illetően a Troodontiákhoz sorolandó, számos hasonlóságot mutatva a Dromaeosauridákkal, de az Archaeopteryx-el is.
Tollazatát tekintve, a felső végtag autopódiumán (=manus, vagyis kéz) 11, a lábszárcsonton 12-13, míg a „csüdön” 10-11 nagy tollra emlékeztető képződmény található. S természetesen, a hosszú farok tájékot is nagyjából hasonló tollak szegélyezik.
A „szárnyon” levő „evezőtollak” vitorlái a Micoraptor-nél és az Archaeopteryx-nél találhatóakkal szemben nem aszimmetrikusak és az I-rendűek hosszabbak, mint a II-rendűek (a kézen lévőket az alkaron elhelyezkedőkhöz viszonyítva). Hasonló tollak találhatóak az alsó végtagon is. A lábszáron levők hosszabbak lévén, mint a lábközépcsonton („csüd”) található s arra merőleges elhelyezkedők. Hasonló, de jóval rövidebb tollak borítják az ujjakat is, egészen a karmokig. Ez is egyedi jelleg és ismeretlen a többi tollas dinoszaurusznál.
Pihe- és pehelyszerű tollak találhatóak a test többi részén, a többi tollas dinoszauruszhoz hasonlóan.
A jellegek alapján a szerzők a leletet az ősi Dromaeosauridákhoz és azoknak is az ősi Troodontia-típusához sorolják. Szerintük az előbbi csoport közös tőről fakadna a madarakhoz vezető Archaeornithes csoporttal. (ld. kladogram)
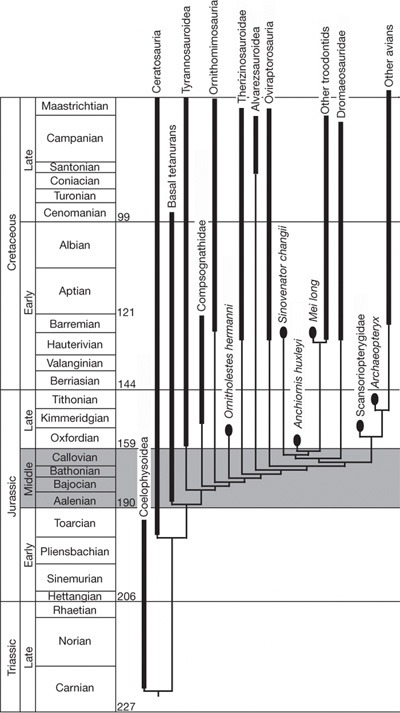
A cikk természetesen nem maradt visszhang nélkül és a folyóirat legújabb számában Lawrence Witmer máris siet kijelenteni, hogy ez a lelet is bizonyítja a madarak dinoszauruszokból való eredetét. Ez természetesen akkor lenne igaz, ha az Archaeopteryx, amelyet a dinoszaurusz-madár eredet hívei konzekvensen madárnak tartanak, tényleg az is lenne. Csak hát, mint már a „Madarak” poszthoz csatolt tanulmányomban kifejtettem, ez egyre kevésbé tűnik valószínűnek.
Mindenesetre, az Anchiornis felfedezése ismét bizonyítja, hogy a teropodák közt nemcsak a Rex-hez hasonló óriás ragadozók, hanem apró termetűek is voltak*, valamint azt is, hogy a tollazat igen hamar megjelent az állatvilágban, ráadásul a hüllők nem is csak egy csoportjánál. Mivel a tollazat funkciói hasonlóak kellett legyenek (védelem, hőszabályozás, költés, ivari kétalakúság biztosítása), nincs miért csodálkozni azon, hogy megjelenési formái sem különböznek jelentősen. Természetesen, lehet következtetni a hosszabb, gerinccel ellátott, evezőtollakhoz hasonló képletekből a repülési képességekre is, de tudomásul kellene venni, hogy azokhoz a tollakon kívül más jellegek létére is szükség van. Valamennyi kistermetű, hosszú tollképletekkel fedett végtagú, dinoszaurusz-típus képes lehetett persze sikló-vitorlázó repülésre, de a valódi madarakra jellemző aktív repüléshez már több kell. De ezeket a csonttani (hiszen a zsigerekre nem hivatkozhatunk, mivel nem maradtak fenn) jellegeket egyetlen tollas dinoszaurusznál, de még az Archaeopteryx-nél sem találjuk meg. Viszont már megvannak a legalsó krétából (144 millió évtől) már ismert valódi madaraknál (Ornithurae-Neornithes).
Mindebből , bárki levonhatja a megfelelő konklúziókat!(* dolphin megj.: a közelmúltban előkerült "mini T-Rex" arra enged következtetni, hogy a Rex-vonal esetében a megnagyobbodott testméret csak később szerzett jelleg.)
Witmer, LM (2009) Palaeontology: Feathered dinosaurs in a tangle. Nature 461: 601-602.
Hu D, Hou L, Zhang L, Xu X (2009) A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus. Nature 461: 640-643.
 A humán paleontológiában nem ritka, hogy egy régi lelet nagy port kavar fel, de ezúttal nemcsak a fosszília kora az, ami miatt a felfedezés "réginek" minősíthető, hanem konkrétan előásásának pillanata is.
A humán paleontológiában nem ritka, hogy egy régi lelet nagy port kavar fel, de ezúttal nemcsak a fosszília kora az, ami miatt a felfedezés "réginek" minősíthető, hanem konkrétan előásásának pillanata is. 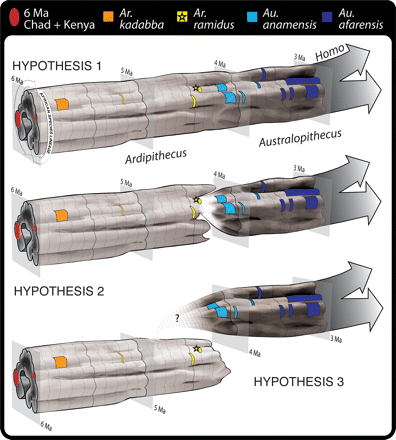 Mit jelent Ardi az "emberré válás" szemszögéből? Nos, drámai bejelentésre ne tessék számítani. Azt már eddig is tudtuk, hogy a
Mit jelent Ardi az "emberré válás" szemszögéből? Nos, drámai bejelentésre ne tessék számítani. Azt már eddig is tudtuk, hogy a  Amit ma a különböző modellorganizmusok fejlődésbiológiájának genetikájáról tudunk, azt elsősorban az elmúlt bő 25 év
Amit ma a különböző modellorganizmusok fejlődésbiológiájának genetikájáról tudunk, azt elsősorban az elmúlt bő 25 év  A csoport legutóbbi cikke, egy másik pikkely-mutánst vesz górcső alá, az ún. "tükör dániót" (
A csoport legutóbbi cikke, egy másik pikkely-mutánst vesz górcső alá, az ún. "tükör dániót" ( Persze ezek alapján jó kérdés, hogy az
Persze ezek alapján jó kérdés, hogy az  Pszeudogénekről, vagyis olyan szekvenciákról, amelyek egykor aktív géneket kódoltak, de mára elvesztették funkciójukat, már
Pszeudogénekről, vagyis olyan szekvenciákról, amelyek egykor aktív géneket kódoltak, de mára elvesztették funkciójukat, már 

Utolsó kommentek